Schlüsselbegriffe: Jahrringanalyse, Klimawandel, Dürre, eingeführte Baumart, Stress, Douglasie, Gemeine Fichte
Abstract
The forest stands of Douglas-fir [Pseudotsuga menziesii (Mirbel) Franco] and Norway spruce [Picea abies (L.) Karst.] in the South Bohemian uplands (Czech Republic) represent an artificial community of a neophyte (native to North America) and indigenous species (with natural distribution in mountain forests). Ecological characteristics of these two tree species and their response to climatic factors were studied using tree ring analyses and correlation of growth indices for tree-to-tree and between trees and historical parameters of climate (average air temperatures, sum of precipitations and relative air humidity). The two datasets were analysed using multidimensional methods. The ordination analysis shows a considerable difference between both tree species. Growth of both species differed significantly by districts and plots. The effect of climate was studied using correlations as floating time-window. Our results showed that average air temperature had a stronger effect on Douglas-fir growth, for which an increment decrease was found at higher temperatures between May 25th and June 24th. Also, the average maximum air temperature affected the growth of Douglas-fir more strongly than that of Norway spruce. More precipitation was associated with a similar positive effect on the growth of both tree species. Air humidity had a significant effect on growth of both studied tree species and the effect of air humidity was more significant than of distinct precipitation. The combined effects of lack of precipitation and high temperature in vegetation period was the major limiting factor for radial growth. Douglas-fir showed higher productivity (+17.2%) and resistance to climate extremes compared to Norway spruce. Our results suggest that a mixed stand could be more resistant in terms of growth stability than mono-specific stands, as the two studied species exhibited different reaction to climate extremes. Our study further showed that mixed stands with high species diversity had a higher production potential (+13.1%).
Zusammenfassung
Die Bestände aus Douglasie [Pseudotsuga menziesii (Mirb.) Franco] und Gemeiner Fichte [Picea abies (L.) Karst.] in den Hochlagen von Südböhmen stellen eine künstliche Gemeinschaft aus einem Neophyten (mit Ursprung in Nordamerika) und einer einheimischer Art (mit natürlicher Verbreitung in Bergwäldern) dar. Die ökologischen Eigenschaften dieser beiden Arten und ihre Reaktion auf klimatische Faktoren wurden hier mithilfe von Jahrringanalysen und der Korrelation von Wachstumsindizes für Baum zu Baum sowie zwischen Bäumen und historischen Klimaparametern (Durchschnittstemperaturen, Summe der Niederschläge und relative Luftfeuchtigkeit) untersucht. Diese zwei Baumarten wurden unter Verwendung mehrdimensionaler Methoden analysiert. Die Ordnungsanalyse zeigte einen erheblichen Unterschied zwischen den beiden Baumarten. Das Wachstum beider Arten unterschied sich je nach Gebiet und Plots erheblich. Die Auswirkung des Klimas wurde unter Verwendung von Korrelationen im gleitenden Zeitfenster untersucht. Die durchschnittliche Lufttemperatur hatte dabei einen stärkeren Einfluss auf das Wachstum von Douglasie, wobei besonders bei höheren Temperaturen von 25. Mai bis 24. Juni ein zunehmender Rückgang des Wachstums festgestellt wurde. Durchschnittliche maximale Lufttemperatur beeinflusste das Wachstum der Douglasie ebenfalls stärker als das Wachstum der Fichte. Höhere Niederschlagsmengen wirkten sich in ähnlichem Maße positiv auf das Wachstum beider Baumarten aus. Die Luftfeuchtigkeit hat auch für das Wachstum beider Baumarten große Bedeutung. Dieser Effekt ist signifikanter als bei ausgeprägten Niederschlägen. Die kombinierten Effekte von Niederschlagsmangel und hoher Temperatur in der Vegetationsperiode war der entscheidende limitierende Faktor für den Radialzuwachs. Douglasie zeigte im Vergleich zu Fichte eine höhere Produktivität (+17,2%) und mehr Resistenz gegenüber Wetterextremen. Ein gemischter Bestand erscheint auf Basis dieser Studie in der Wachstumsstabilität widerstandsfähiger als Reinbestände, da beide Arten unterschiedlich auf Wetterextreme reagieren. Darüber hinaus zeigten unsere Ergebnisse, dass gemischte artenreiche Waldbestände ein höheres Produktionspotential (+13,1%) aufweisen.
1. Introduction
Douglas-fir (Pseudotsuga menziesii (Mirbel) Franco) is one of the most widespread commercially used tree species not only in the area of its native range, but also in many countries on other continents (Hermann and Lavender, 1999; Podrázský and Kupka, 2011; Kubeček et al., 2014; Mondek and Baláš, 2019). It has become one of the most commercially important introduced tree species in Europe (Essl, 2005; Schmid et al., 2014; Ulbrichová et al., 2014), thanks to its high wood production, adaptability, and wide ecological niche within a large part of the spectrum of European natural conditions. As of 2018, it is growing on an area of 0.83 mil. ha in Europe (Brus et al., 2019). The extent of suitable sites may even extend in future, particularly on mountain and mountain-foothills sites (Pötzelsberger et al., 2019). Douglas-fir is also a potentially important introduced tree species in the forestry sector of the Czech Republic and its importance will increase in the coming years (Kubeček et al., 2014; Slodičák et al., 2014; Vašíček, 2014; Novák et al., 2018; Mondek and Baláš, 2019). Heat and drought waves are expected to become more frequent and intense in our latitude in the future (Schär et al., 2004; Giorgi and Coppola, 2007; Macků and Kosová, 2020), and a number of adaptation measures are needed, including the use of introduced tree species and the so-called assisted migration (Nadezhdina et al., 2014; Gömöry et al., 2020) to a lesser extent.
Douglas-fir is considered as tree species resistant to climatic extremes (Vitali et al., 2017). Damages by early and late frosts that occur commonly in the Central European area (Gallo et al., 2014; Gallo et al., 2017), to which particularly young stands are sensitive (Šindelář, 2003; Chakraborty et al., 2019), and also susceptibility to winter over-transpiration, the so-called physiological drought (Hofman, 1964), are an exception. Douglas-fir growth correlates with climate, soil moisture regime, and soil nutrient status (Eckhart et al., 2019). It has been demonstrated that decreased water availability limits its growth more than the temperature or length of the growing season (Littell et al., 2008) in its natural range. On the other hand, lower success is shown on heavy soils with higher water content (Perić et al., 2011). A significant increment reduction and signs of decline associated with soil moisture deficit were also evidenced by the study of Sergent et al. (2014). Douglas-fir is more resistant to drought than Norway spruce (Picea abies L. Karst) (Nadezhdina et al., 2014) and can also represent an important stabilizing element of forest stands, as reported by Sergent et al. (2010) in study from France, thanks to its powerful root system (Ulbrichová et al., 2014; Mauer and Vaněk, 2014), which is rooting even in deeper soil layers (Mauer and Palátová, 2012). According to Urban et al. (2011), Douglas-fir is resistant to drought in younger stands and prolonged drought in mature stands. Douglas-fir draws water from deeper soil layers much more efficiently than Norway spruce, transposing more water during dry periods (Šach et al., 2019).
Douglas-fir also has a positive effect on habitats compared to Norway spruce (Podrázský and Remeš, 2008; Menšík et al., 2009; Kacálek et al., 2010; Kupka et al., 2013; Ulbrichová et al., 2014), its effect was also positive on afforested agricultural land (Kupka and Podrázský, 2010). This tree species also has a favourable effect on the state of forest phytocoenoses, compared to Norway spruce (Viewegh et al., 2014; Matějka et al., 2015). Nevertheless, it is hypothesized that there is a danger of invasiveness in future (in case of rapid change of environmental conditions) connected with possible intensive natural regeneration of Douglas-fir and disbalancing of nutrient (nitrogen) regime under conditions of high-level nitrogen depositions (Podrázský et al., 2014b; Matějka et al., 2015; Podrázský et al., 2020). Therefore, as with other introduced tree species, it is recommended to cultivate it carefully and with respect to the nature conservation interests (Kuneš et al., 2019; Brundu et al., 2020). Currently, Douglas-fir does not show an invasive character in Europe (Pötzelsberger et al., 2020), but it can be successfully cultivated from natural regeneration under appropriate silvicultural measures (Eberhard and Hasenauer, 2018).
Considering its advantages and possible limitations, some authors currently propose Douglas-fir as possible alternative to Norway spruce on the habitats corresponding to the managed forests of Central Europe (e.g. Sergent et al., 2010; Fischer and Neuwirth, 2012; Kubeček et al., 2014; Podrázský, 2016; Podrázský et al., 2016; Vitali et al., 2017; Novák et al., 2019; Remeš et al., 2020). In the future, in connection with global climate change, the use of Douglas fir in the Czech Republic is expected on fresh habitats, where it could replace the economic production of Norway spruce to some extent. Under the conditions of the Czech Republic, the Douglas fir acts similarly to silver fir (Abies alba Mill.) in terms of habitats and especially vegetation conditions (Viewegh et al., 2014). Douglas fir is planned for use mainly in mixtures with European beech, common and pedunculate oak, sycamore maple, silver fir and European larch (Poleno et al., 2009). In the Czech Republic, Douglas fir is not listed in the list of invasive tree species (Pyšek et al., 2012; Pergl et al., 2016). In the forestry of the Czech Republic, Douglas fir is used as a stabilising and ameliorative tree, but only in a limited number, as it is a non-genetically native species, and the legislation does not allow the full use of its potential (Mlíkovský and Stýblo, 2006). This strategy is based on the National Forestry Program II and the Principles of the State Forestry Policy, which were approved by the Government of the Czech Republic (MZE, 2013). At present, there are over 6,000 hectares of Douglas fir in the Czech Republic, which represents 0.24% in the species composition of forests. Over the last ten years, area of Douglas fir stands has grown at an average of 120 hectares per year (MZE, 2020). It can thus help mitigate the expected decline in the supply of softwood in the near future (Podrázský et al., 2014a; Pulkrab et al., 2015; Palátová et al., 2017) and mitigate the effects of the current situation within the forestry sector in the Czech Republic (Riedl et al., 2019). The cultivation in mixed stands with indigenous tree species including broadleaves like European beech (Podrázský et al., 2015) or birches including rare taxons (e.g. Baláš et al., 2016) and the selection of suitable provenances (Kšír et al., 2015) are emphasised for Douglas-fir silviculture, as the stability of forest stands is increased with higher number of tree species (Vacek et al., 2020a). Its potential to elevate mobile soil nitrogen levels can be significantly limited in this way (Zeller et al., 2010; Podrázský et al., 2014b; Podrázský et al., 2020). Regarding provenances, researches from Bulgaria suggest that Douglas-fir of continental origin is more susceptible to fungal pathogens Phaeocryptopus gaeumannii (Rohde) Petrak and Rhabdocline pseudotsugae (Syd.) in comparison to Western Cascade Mountains and Coastal-originated Douglas-firs that are characterised by more rapid growth and higher resistance to those pathogens (Petkova et al., 2014). Extremely high productivity was found in case of such suitable provenances (Popov, 2014). However, characteristics of microsites need to be considered for optimal production, as reported by Čater (2021), in a study from Slovenia. Growth response is highly dependent on the site index and competition intensity (Castaldi et al., 2017). High-quality Douglas-fir stands with unknown origin can be tracked back by assignment using Bayesian cluster analysis to determine the origin of seeds (Hintsteiner et al., 2018). It was found out that a bottleneck effect occurs for next generation of Douglas-fir stands in terms of genetic diversity in Central Europe, which needs to be addressed in careful management (Neophytou et al., 2020). Seed lots from original provenances in North America should be preferred over seeds from Europe when establishing new plantations in Europe for the same reason (Eckhart et al., 2017). Both coastal and interior varieties, and their hybrids are present in European forests (van Loo et al., 2019).
It seems useful to know the difference in growth between Norway spruce and Douglas-fir under comparable conditions. Norway spruce growth at lower and medium altitudes is strongly endangered by the ongoing climate change (Vacek et al., 2019), or this tree species should not be cultivated on non-wet soils to approximately 4th forest vegetation zone, and that Douglas-fir is non-indigenous species in Europe (Carrillo-Gavilán et al., 2012; Tschopp et al., 2015).
Recent discussions are related to diversity and productivity of Douglas-fir stands (Kupka et al., 2013; Podrázský et al., 2013). Available scientific results regarding Douglas-fir from Central European area include its seed performance (Kuneš et al., 2017), effect on grass/herbaceous communities, which were studied e.g. as artificial mixtures in pot experiments (Špačková and Lepš, 2001), fertilization experiments (Cukor et al., 2017a; Vacek et al., 2019b), growth and survival dynamics on abandoned agricultural lands (Hedlund et al., 2003; Cukor et al., 2017b), and carbon sequestration (Podrázský et al., 2018). Diversity and productivity of stands with Norway spruce are studied in forests (Vacek, 2017; Vacek et al., 2019c) where these relations would be influenced by the difference in traits of the tree species of the stand (e.g. Körner, 2005; Vacek et al., 2021). The effect of diversity on production can arise from different allocation of resources by different species and/or from different stress resistance related to various growth depression. The mixture of both studied species under examination in the South Bohemia uplands represents an artificial community of the neophyte (Pseudotsuga menziesii originated in North America) and species with natural distribution dominantly in higher vegetation zones (Picea abies from mountain forests). The aim of the presented study was to look for potential links between climatic factors (air temperature, precipitation, air humidity) and the radial growth of Douglas-fir and Norway spruce, for which correlation analyses are used.
2. Materials and Methods

Figure 1: Location of the research region in the Czech Republic and location of four districts of research plots (Vodňany, Kamýk, Vráž and Sedlice) together with the main meteorological stations of CHMI (Kocelovice, Nadějkov, Vráž, Strakonice and Temelín); the gray area in map shows the forest cover and the gray lines the Natural Forest Area. / Abbildung 1: Lage der Forschungsregion in der Tschechischen Republik und Lage von vier Bezirken von Forschungsflächen (Vodňany, Kamýk, Vráž und Sedlice) zusammen mit den wichtigsten Wetterstationen des CHMI (Kocelovice, Nadějkov, Vráž, Strakonice und Temelín); die Grauzone auf der Karte zeigt die Waldbedeckung und die grauen Linien die Naturwaldgebiet.
2.1 Study area
The research took place in the Southern Bohemia region, in the Písek district, in four areas with mature commercial forests with a different representation of Picea abies and Pseudotsuga menziesii (Fig. 1). The study area is located at an altitude of 455–587 m asl with an average slope of 5 degrees. Cambisols and Luvisols are prevailing soils types (Němeček et al., 2001). The bedrock is mainly composed of gneiss, granite and migmatite. The annual average air temperature is 8.7 °C with a maximum monthly mean in July (18.6 °C) a minimum monthly mean in January (-1.0 °C) in the period 1989–2019. The annual sum of precipitation reaches 610 mm, while the highest sum is in July (89 mm) and the lowest in February (22 mm). The average number of days with snow cover is 49. The growing season (Tmax > 10 °C) lasted approximately 154 days, with a mean precipitation of 415 mm and a mean temperature of 14.9 °C. The number of tropical days (Tmax > 30 °C) was 9, the number of ice days (Tmax < 0 °C) was 30, and the number of arctic days (Tmax < -10 °C) was 0. The study territory has typically warm and dry summers and cool and dry winters with a narrow annual temperature range (Cfb) according to Köppen climate classification (Köppen, 1936).

Table 1: Summary of research plots with Pseudotsuga menziesii (Pm) and Picea abies (Pa) (dominant and subdominant trees are marked in bold). Lines in italics represent plots on which no cores were taken. / Tabelle 1: Zusammenfassung der Forschungsflächen mit Pseudotsuga menziesii und Picea abies (dominante und subdominante Bäume sind fett markiert). Kursiv gedruckte Linien stellen Forschungsgebiete dar, auf denen keine Bohrungen vorgenommen wurden.
A single-story forest stands with a minimum age of 80 years and a well-developed phytocoenoses were selected for the study. The stands belong predominantly to a eutrophic series of fresh, nutrient-medium category of Querceto-Fagetum mesotrophicum (Viewegh et al., 2003). Sub-xerophile ecotype of common oak (Quercus robur L.) and European beech (Fagus sylvatica L.) should be the dominant species in the potential reconstructed natural tree composition at all plots, however, at present, these species form only an admixture with a representation of up to 5% in the monitored stands. Other scattered and admixed tree species occurring in the research areas include Scots pine (Pinus sylvestris L.), larch (Larix decidua Mill.), silver fir (Abies alba Mill.), European beech (Fagus sylvatica L.) and silver birch (Betula pendula Roth). The list of plots and their characteristics are summarised in Table 1.
2.2 Data collection
A total of 25 research plots with a radius of 12.62 m (size 500 m2) were established at the beginning of autumn 2019. The coordinates of their centres were determined on all plots using a Trimble Geo 7X instrument with TerraSync software. The positions of all trees were measured on the plots, the species was recorded, the diameter at the breast height (DBH), the height of the tree, the height of the crown base, the crown projections were measured and the tree injury was recorded using the inventory Field-Map system (IFER-Monitoring and Mapping Solutions Ltd.). All individuals of the tree layer with a DBH ≥ 4 cm were recorded. The DBH was measured by Mantax Blue metal caliper (Haglöf, Sweden) with an accuracy of 1 mm, and tree height was measured by Laser Vertex hypsometer (Haglöf, Sweden) with an accuracy of 0.1 m. A total of 14 plots were selected for tree-ring analysis. These plots were selected on the basis of the following ratio of spruce and Douglas fir: 1) <10% – individually mixed tree species; 2) 10–29% – mixed and scattered species, 3) 30–49% – basic species, 4) 50–89% – predominant species, 5) 90–100% – monoculture. A minimum of 22 cores (maximum 26) were taken from both tree species from each variant, always from two localities. Increment cores were taken from 247 trees (121 Picea abies and 126 Pseudotsuga menziesii). In most cases, core samples were taken directly on plot or in immediate proximity (maximum 20 m from the plot) in case of not enough suitable trees growing directly on the plot due to low proportion of examined tree species. Only codominant and dominant trees according to the classification by Kraft (1884) were chosen as the significant growth response (compared to subdominant and suppressed trees; Remeš et al., 2015). These trees were randomly (RNG in Excel) selected for a dendrochronology analysis. Individual increment cores were taken by Presler's increment borer (Mora, Sweden) up to the centre of the tree or to a depth of 30 cm at least in autumn of 2019. The increment cores were taken not only on the research areas, but also in close proximity, to assure more samples. Annual ring widths were measured with an accuracy of 0.01 mm by an Olympus binocular microscope (Olympus, Japan) on the LINTAB measuring table (Rinntech, Germany) and recorded with TsapWin software (Resistograph, Germany).
2.3 Data analysis
The tree-ring increment series were individually crossdated (to remove errors caused by missing tree rings) using statistical tests in the PAST application (Knibbe, 2007) and subsequently they were subjected to visual inspection according to Yamaguchi (1991). If a missing tree ring was revealed, a tree ring of 0.01 mm in width was inserted in its place. Individual tree ring curves from each tree were age-detrended and an average tree-ring series was created in the ARSTAN programme (Cook, Tree Ring Laboratory). A negative exponential spline and subsequently 0.67n spline were used for the age-detrending (Grissino-Mayer et al., 1992). This process results in a series of yearly growth indices for each tree.
The analysis of negative pointer years was done according to Schweingruber (1996) and Desplanque et al. (1999). For each tree the pointer year was tested as an extremely narrow tree ring that does not reach 40% of the increment average from the four preceding years. The occurrence of the negative year was proved if such a strong reduction in increment occurred at least in 20% of trees on the plot.
Growth index data between 1965 and 2019 were chosen for further analysis, because this period is covered by data for all trees except for the 3 youngest individuals, whose data were excluded from processing. Growth index curves were correlated (Pearson's linear correlation coefficient r was used) for all pairs of individuals. These correlation coefficients form a symmetric correlation matrix. This matrix was used for ordination analysis by PCA method (Statistica program was used for calculation, graphs were created in PlotOA program). Only these correlations are input variables for PCA. The PCA results were interpreted in terms of description of the growth variability among trees representing both species (differences of both species) and different localities (differences among localities). Two trees which are localized near one to the other in the ordination graph shows more synchronized growth comparing a pair of trees with higher distance in the ordination graph. Results of PCA are presented as graphs of the first two PCA axes. Distribution ellipses embracing 95% of the objects (trees) of the category (species, locality etc.) are used to elucidate difference among different species (localities).
The correlation matrix was used to calculate the distance matrix, where dij = 1 - rij, at the same time. This distances matrix was used for cluster analysis (classification of individuals) by the Ward's method (the Cluster program from the DBreleve package was used – Matějka, 2020).
Five meteorological stations of the Czech Hydrometeorological Institute (www.chmi.cz) are in the wider interest district. However, only the Vráž station (indicative C1VRAZ01), located close to the research areas, has the highest potential for use, i.e. at a similar altitude and, moreover, in a landscape with a high proportion of forests.
Climatic characteristics from the Vráž station were evaluated using a time moving window (interval) with variable length (count of days) and position (described by the last day of the interval as order number of the day within year [DoY - day of year]). This two-dimensional application of floating window arises from methods e.g. astrochron in the R package (https://rdrr.io/cran/astrochron/man/mwCor.html) or in Sageman and Hollander (1999). The average climate characteristics for this year were calculated for each year (with a possible overlap to the previous year) due to this window. The correlation coefficient between the average value for the climate within the floating window in each year and the radial increment index in the corresponding year for the individual trees of both species was calculated in the next step. The set of the resulting correlation coefficients was compiled into graphs, from which the position of the extreme values of the correlation coefficient is evident. Position of the moving time window is defined by the end day of the window as the horizontal coordinate and width of the window as number of days equal to the vertical coordinate. This makes it possible to estimate in which time interval the given climatic characteristic has the greatest influence on the growth of the given tree species.
3. Results

Table 2: Species composition based on the tree basal area (m2·ha−1) according to the individual plots of size 500 m2. / Tabelle 2: Artenzusammensetzung basierend auf der Baumgrundfläche (m2·ha−1) gemäß den einzelnen Parzellen mit einer Größe von 500 m2.
Only a small number of tree species was present in the research plots. One of both species was dominant or subdominant at least. The total tree canopy cover was evaluated between 40 and 85 %. (Table 1). The tree basal area varies between 41.1 and 85.2 m2·ha−1 (Table 2). The mean basal area in dominant Picea abies stands was 54.2 m2·ha−1, while it was by 17.2% more in Pseudotsuga menziesii (63.5 m2·ha−1). Species-rich stands with an admixture of more than two deciduous tree species (m = 4+) showed a higher basal area by 13.1%.
The variability of the average growth curves is considerable for both species. The classification of these curves using cluster analysis is suitable (Fig. 2). Individual clusters can be distinguished according to which tree species predominates in the cluster.

Figure 2: Increment cores classification by Ward’s method. Clusters with a Picea abies dominance are in black, clusters with a Pseudotsuga menziesii are in red (blue line – clustering level used for definition of the descripted clusters). The vertical axis – dissimilarities based on correlations, see methods. / Abbildung 2: Klassifizierung der Bohrungen nach der Ward-Methode Cluster mit einer Picea abies – Dominanz sind schwarz, Cluster mit einer Pseudotsuga menziesii sind rot (blaue Linie – Clustering-Ebene, die zur Definition der beschriebenen Cluster verwendet wird). Die vertikale Achse – Unähnlichkeiten aufgrund von Korrelationen, siehe Methoden.

Figure 3: Year-on-year variability of Picea abies diameter increment in individual main classification groups of individuals according to Figure 2. The numbers in the box are binary codes for the designation of classification groups, which are based on the branching of the binary tree graph of the classification. / Abbildung 3: Die Variabilität von Picea abies gegenüber dem Vorjahr in einzelnen Hauptklassifikationsgruppen von Individuen gegenüber dem Vorjahr. Die Zahlen in der Box sind Binärcodes für die Bezeichnung von Klassifikationsgruppen, die auf der Verzweigung des Binärbaumgraphen der Klassifikation basieren.
The maximum Picea abies increment was recorded in 1967, 1985, 1988–9, 1997, 2002 and 2011 (really in the whole period 2009–2014). A decrease in increment occurred in 1976, 1993, 2000, 2007, 2015 and 2018–2019 on the contrary (Fig. 3). In 1993, the lack of precipitation was observed at the begging of the vegetation period, i.e. the driest March (8 mm, mean 36 mm) from the period 1989 – 2019. Negative pointer years 2015, 2018 and 2019 were characterized by extremely high temperature during the vegetation period with below-average precipitation. Historically, the highest number of tropical days (29 days with Tmax ≥ 30 °C) was recorded in 2015. The warmest April (13.5 °C, mean 8.7 °C) and May (16.1 °C, mean 13.1 °C) was observed in 2018, respectively the warmest June (21.4 °C, mean 16.7 °C) in 2019.

Figure 4: Year-on-year variability of Pseudotsuga menziesii diameter increment in individual main classification groups of individuals according to Figure 2. The numbers in the box are binary codes for the designation of classification groups, which are based on the branching of the binary tree graph of the classification. / Abbildung 4: Variabilität des Durchmesserinkrements von Pseudotsuga menziesii in einzelnen Hauptklassifkationsgruppen von Individuen gegenüber dem Vorjahr. Die Zahlen in der Box sind Binärcodes für die Bezeichnung von Klassifikationsgruppen, die auf der Verzweigung des Binärbaumgraphen der Klassifikation basieren.
The situation was different for Pseudotsuga menziesii. The maximum values were recorded in 1966, 1997, 2002, 2009 and 2014, while the minimums were recorded in 1976, 1979, 2013 and 2018; an only slight decrease was in 2003 (Fig. 4). Synergism of the warmest (22.3 °C, mean 18.4 °C) and driest August (22 mm, mean 79 mm) was observed in 2003. The historically warmest day was recorded within this year: 13 August (36.8 °C). The year 2013 was characterized by an extremely high sum of precipitation during the vegetation period (higher by 135%) and cloudy weather (low sunshine). Regarding the year 2018, see description in Picea abies. Generally, comparing pointer years with extremely low or high radial growth in both species, only 2 same pointer years were found for maximum (1997 and 2002) and 2 for minimum (1976 and 2018).

Figure 5: Position of Picea abies (SM) and Pseudotsuga menziesii (DG) individuals in the PCA ordination space based on correlations of their growth. / Abbildung 5: Position von den Individuen Picea abies (SM) und Pseudotsuga menziesii (DG) im PCA-Ordnungsraum basierend auf Korrelationen ihres Wachstums.
The first PCA ordination axis represents 79.0 % of the total data variance, the second axis is corresponding to 7.9 % of the total variance. The ordination analysis shows a considerable difference between both tree species, which is manifested by the position of the respective points in the ordination space and thus also by the mutual displacement of the distribution ellipses (Fig. 5).
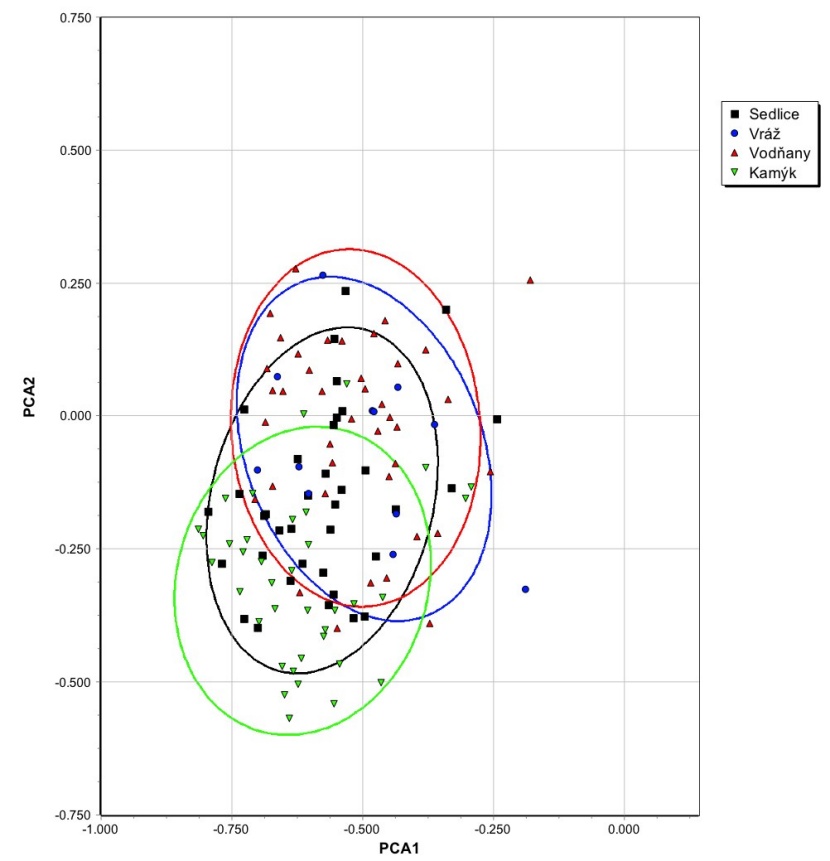
Figure 6: Position of Pseudotsuga menziesii individuals in the PCA ordination space based on correlations of their growth. The tree species affiliation to the district is highlighted and 95% distribution ellipses are drawn. / Abbildung 6: Position von Pseudotsuga-menziesii-Individuen im PCA-Ordnungsraum basierend auf Korrelationen ihres Wachstums. Die Zugehörigkeit der Baumarten zum Distrikt wird hervorgehoben und es werden 95%ige Verteilungsellipsen gezeichnet.
Pseudotsuga menziesii growth differs by districts and plots considerably. The growth dynamics of Pseudotsuga menziesii is similar to the growth dynamics of Picea abies on plot 14 (district Vodňany), but the results are significantly different on plots 21 and 24 (district Kamýk) especially. The influence of the locality on the growth dynamics of both species is significant (Fig. 6 and 7), while the importance of the locality is higher in Picea abies (distribution ellipses are less overlapped) than in Pseudotsuga menziesii (distribution ellipses are more overlapped), which may indicate that Picea abies is a species growing in Písek region in unsatisfactory environmental conditions.

Figure 7: Position of Picea abies individuals in the PCA ordination space based on correlations of their growth. The tree species affiliation to the district is highlighted and 95% distribution ellipses are drawn. / Abbildung 7: Position von Picea-abies-Individuen im PCA-Ordnungsraum basierend auf Korrelationen ihres Wachstums. Die Zugehörigkeit der Baumarten zum Distrikt wird hervorgehoben und es werden 95%ige Verteilungsellipsen gezeichnet.
The climate was changing during the observation period (1961–2019) in the region. The data from the meteorological station Vráž show a significant increase in the yearly average air temperature (+0.029 °C per year, probability P near 100.0 %), similarly to yearly absolute minima (+0.050 °C per year, P = 94.5%) and maxima (+0.035, P = 99.0 %). The yearly sum of precipitations was strongly variable without any trend. A similar situation is with the average relative air humidity, where minimal averages were recorded in 1973 (72.5%), 1986 (73.0%; the humidity depression was in the whole period 1982–1990) and 2015 (73.7%). On the other hand, the maximum average was in 2001 (82.7%). The period 2001–2019 can be characterized by the significant trend of decrease in air humidity (-0.34% per year).

Table 3: The extreme values of correlation coefficients (r) between climate parameters within variable interval (position specified by end day of year and length of the interval) and index of diameter growth. DoY – day of year. See figs. 8–12. All values with |r| > 0.004 are significant at level α = 0.05. n – number of cases (pairs of values weather parameter and index of diameter growth). / Tabelle 3: Die Extremwerte der Korrelationskoeffizienten (r) zwischen Klimaparametern innerhalb eines variablen Intervalls (Position angegeben durch den Endtag des Jahres und die Länge des Intervalls) und den Index des Durchmesserwachstums. DoY – Tag des Jahres. Siehe Abb. 8–12. Alle Werte mit |r| > 0,004 sind bei Stufe α = 0,05 signifikant. n – Anzahl der Fälle (Wertepaare Wetterparameter und Index des Durchmesserwachstums).
Correlation among climatic features and growth of both species using floating window are drawn in Figs. 8–12 and in Table 3. The average air temperature has a stronger effect on Douglas-fir growth, for which an increment decrease was found at higher temperatures between May 25th and June 24th. The growth is positively affected by higher temperatures between the beginning of February and April 10th, which is usually associated with an earlier start of growth. High average temperatures from mid-July to mid-August are associated with reduced increment for Norway spruce – due to the optimum Norway spruce occurrence at higher altitudes, this tree species suffers from high summer temperatures at Písek region (Fig. 8).

Figure 8: Correlation between average air temperature calculated within floating window (interval of specified end day on the horizontal axis and length along the vertical axis) and index of radial growth by Picea abies (left; minimal r = -0.23, maximal r = 0.07) and Pseudotsuga menziesii (right; minimal r = -0.20, maximal r = 0.23) during 1961–2019. Correlation coefficient is marked by size of the point with position corresponding to the order-number of end day within year and length of the floating window (colour pink if r < 0 and blue if r > 0), see Methods and Table 3. / Abbildung 8: Korrelation zwischen der durchschnittlichen Lufttemperatur innerhalb des Schwimmfensters (Intervall des angegebenen Endtages auf der horizontalen Achse und der Länge entlang der vertikalen Achse) und dem Index des radialen Wachstums durch Picea abies (links; minimal r = -0,23, maximal r = 0,07) und Pseudotsuga menziesii (rechts; minimal r = -0,20, maximal r = 0,23) im Zeitraum 1961–2019. Der Korrelationskoeffizient wird durch die Größe des Punktes mit der Position markiert, die der Bestellnummer des Endtages innerhalb des Jahres und der Länge des schwebenden Fensters entspricht (Farbe rosa, wenn r < 0 und blau, wenn r > 0), siehe Methoden und Tabelle 3.
The minimum air temperatures were less related to Norway spruce growth in comparison to Douglas-fir. The positive effect of higher minimum air temperatures on Norway spruce growth was indicated in two periods - from mid-March to mid-April and from mid-June to mid-July. In contrast, Norway spruce growth depression is correlated with low minimum temperatures in winter (approximately from November to January), which may be related to the tree species frost damage and this is probably accentuated by growth in an area with often insufficient snow cover. Higher minimum air temperatures from mid-March to mid-April are associated with an increase in Douglas-fir growth, which clearly indicates the importance of the early beginning of the growing season (Fig. 9).

Figure 9: Correlation between average daily minimum of air temperature calculated within floating window (interval of specified end day on the horizontal axis and length along the vertical axis) and index of radial growth by Picea abies (left; minimal r = -0.12, maximal r = 0.11) and Pseudotsuga menziesii (right; minimal r = -0.15, maximal r = 0.25) during 1961–2019. Correlation coefficient is marked by size of the point with position corresponding to the order-number of end day within year and length of the floating window (colour pink if r < 0 and blue if r > 0), see Methods and Table 3. / Abbildung 9: Korrelation zwischen dem durchschnittlichen täglichen Minimum der Lufttemperatur, berechnet innerhalb des Schwimmfensters (Intervall des angegebenen Endtages auf der horizontalen Achse und der Länge entlang der vertikalen Achse) und dem Index des radialen Wachstums durch Picea abies (links; minimal r = -0,12, maximal r = 0,11) und Pseudotsuga menziesii (rechts; minimal r = -0,15, maximal r = 0,25) im Zeitraum 1961–2019. Der Korrelationskoeffizient wird durch die Größe des Punktes mit der Position markiert, die der Bestellnummer des Endtages innerhalb des Jahres und der Länge des schwebenden Fensters entspricht (Farbe rosa, wenn r < 0 und blau, wenn r > 0), siehe Methoden und Tabelle 3.
The average of the maximum air temperatures again affects the growth of Douglas-fir more than the Norway spruce growth. Norway spruce shows the growth depression occurrence at high maximum temperatures. This relates to the occurrence of high temperatures in June (the period of the most intensive growth) and to mid-August (high temperatures probably end a radial growth prematurely) in the current year. The negative impact of maximum temperatures high averages during the long period from July of the previous year to January of the current year is even more significant - such high temperatures probably cause stress, which the tree species must cope in the following year. In the case of Douglas-fir, the most significant positive effect is the high average daily maximum air temperatures in the early spring period from February to the first half of April, which again indicates the effect of the growing season beginning (Fig. 10).

Figure 10: Correlation between average daily maximum of air temperature calculated within floating window (interval of specified end day on the horizontal axis and length along the vertical axis) and index of radial growth by Picea abies (left; minimal r = -0.23, maximal r = 0.07) and Pseudotsuga menziesii (right; minimal r = -0.18, maximal r = 0.22) during 1961–2019. Correlation coefficient is marked by size of the point with position corresponding to the order-number of end day within year and length of the floating window (color pink if r < 0 and blue if r > 0), see Methods and Table 3. / Abbildung 10: Korrelation zwischen dem durchschnittlichen täglichen Maximum der Lufttemperatur, berechnet innerhalb des Schwimmfensters (Intervall des angegebenen Endtages auf der horizontalen Achse und der Länge entlang der vertikalen Achse) und dem Index des radialen Wachstums durch Picea abies (links; minimal r = -0,23, maximal r = 0,07) und Pseudotsuga menziesii (rechts; minimal r = -0,18, maximal r = 0,22) im Zeitraum 1961–2019. Der Korrelationskoeffizient wird durch die Größe des Punktes mit der Position markiert, die der Bestellnummer des Endtages innerhalb des Jahres und der Länge des schwebenden Fensters entspricht (Farbe rosa, wenn r <0 und blau, wenn r > 0), siehe Methoden und Tabelle 3.
Higher totals of precipitation are associated with a positive effect on the growth of both tree species to a similar extent. It is necessary to evaluate the total precipitation for the entire vegetation period (until the first half of August) for Norway spruce. The significance of sufficient precipitation at the beginning of the growing season (in April) is only very weakly indicated. The most significant is the high total precipitation in July, for Douglas-fir. The importance of sufficient precipitation in April and in the first part of May is also indicated (Fig. 11).

Figure 11: Correlation between total rain calculated within floating window (interval of specified end day on the horizontal axis and length along the vertical axis) and index of radial growth by Picea abies (left; minimal r = -0.09, maximal r = 0.30) and Pseudotsuga menziesii (right; minimal r = -0.14, maximal r = 0.29) during 1961–2019. Correlation coefficient is marked by size of the point with position corresponding to the order-number of end day within year and length of the floating window (color pink if r < 0 and blue if r > 0), see Methods and Table 3. / Abbildung 11: Korrelation zwischen dem im schwimmenden Fenster berechneten Gesamtregen (Intervall des angegebenen Endtages auf der horizontalen Achse und der Länge entlang der vertikalen Achse) und dem Index des radialen Wachstums durch Picea abies (links; minimal r = -0,09, maximal r = 0,30) und Pseudotsuga menziesii (rechts; minimal r = -0,14, maximal r = 0,29) im Zeitraum 1961–2019. Der Korrelationskoeffizient wird durch die Größe des Punktes mit der Position markiert, die der Bestellnummer des Endtages innerhalb des Jahres und der Länge des schwebenden Fensters entspricht (Farbe rosa, wenn r < 0 und blau, wenn r > 0), siehe Methoden und Tabelle 3.
The influence of air humidity on their growth is strongly significant for both tree species. This effect is more significant than in the case of distinct precipitation. The high air humidity in the second half of the summer (for 2 to 3 months) has a positive effect, the same for both species (Fig. 12). Higher air humidity in this period indicates sufficient total precipitation, but also more even rainfall distribution. A higher total precipitation can be achieved by short-term intense rain, a large part of which drains off and does not penetrate into the soil. However, such torrential rain will not cause a more permanent increase in air humidity. Therefore, air humidity should be evaluated regularly as part of climate analyses (and not only in relation to tree species growth).

Figure 12: Correlation between average relative air humidity calculated within floating window (interval of specified end day on the horizontal axis and length along the vertical axis) and index of radial growth by Picea abies (left; minimal r = -0.11, maximal r = 0.32) and Pseudotsuga menziesii (right; minimal r = -0.17, maximal r = 0.30) during 1961–2019. Correlation coefficient is marked by size of the point with position corresponding to the order-number of end day within year and length of the floating window (color pink if r < 0 and blue if r > 0), see Methods and Table 3. / Abbildung 11: Korrelation zwischen der durchschnittlichen relativen Luftfeuchtigkeit, die innerhalb des Schwimmfensters berechnet wurde (Intervall des angegebenen Endtages auf der horizontalen Achse und Länge entlang der vertikalen Achse) und dem Index des radialen Wachstums durch Picea abies (links; minimal r = -0,11, maximal r = 0,32) und Pseudotsuga menziesii (rechts; minimal r = -0,17, maximal r = 0,30) im Zeitraum 1961–2019. Der Korrelationskoeffizient wird durch die Größe des Punktes mit der Position markiert, die der Bestellnummer des Endtages innerhalb des Jahres und der Länge des schwebenden Fensters entspricht (Farbe rosa, wenn r < 0 und blau, wenn r > 0), siehe Methoden und Tabelle 3.
4. Discussion
Douglas fir in natural conditions on the American continent is characterized by great variability in the morphological and physiological features of its two main ecotypes: the Rocky Mountain (mountain) ecotype and the Pacific coastal ecotype (Harlow et al., 1979). These variations also occur in populations introduced to Europe. In addition to morphological differences, this also applies to growth rate, resistance to wind, frost, drought and seasonal diversification of growth rhythm (Chylarecki, 1976; Feliksik and Wilczyński, 2004; Wilczyński and Feliksik, 2007). Douglas fir thus points to a wide range of reactions to environmental conditions and its individual plasticity (Schober, 1963; Białobok and Mejnartowicz, 1970; Mejnartowicz, 1976; Wilczyński and Feliksik, 2007), as in our case.
Both Pseudotsuga menziesii and Picea abies in the studied region show a strong influence of the locality. It is valid especially for Picea abies, which indicates that it grows in unsuitable conditions here and its growth in these altitudes is strongly endangered; as mentioned in other studies, too (Vacek et al., 2019a). Both species are influenced by climate in different manner. Precipitation, temperature and extreme climatic events, for example droughts, are strongly reflected in radial growth of Douglas-fir (Littell et al., 2008; Arreola-Ortiz et al., 2010; Sergent et al., 2014; Lachenbruch and Johnson, 2020) and Norway spruce (Mikulenka et al., 2020; Vacek et al., 2020c). The significant effect of air temperatures and the sum of precipitation, especially their distribution during the year also play an important role (Acosta-Hernández et al., 2017; Gallo et al., 2020a).
The high average air temperatures in summer are associated with reduced growth for Picea abies. Similarly, negative effect of air temperature on spruce radial growth was observed in the period from May to July in lowland forests in another part of the Czech Republic (Vacek et al., 2019a). The opposite situation was observed in spruce mountain forests, where low temperature was the limiting factor of spruce growth (Král et al., 2015; Cukor et al., 2020; Vacek et al., 2020b). Generally, the limiting effect of low temperatures was more significant at high-altitude sites, while the importance of precipitation increased at low altitudes (Mäkinen et al., 2002). Moreover, the high maximum air temperatures have negative effect until January in our study. The tree species cope with this stress in the following year. Minimum air temperatures are only minimally related to spruce growth conversely. Higher sum of precipitation with even distribution throughout the vegetation period is associated with a positive effect.
The high average air temperatures during June correlate with an increment decrease for Pseudotsuga menziesii. A similar conclusion is reached by the study of Wilczyński and Feliksik (2007) and Eckhart et al. (2019), in which they also report a lower production of Douglas-fir in the regions with the highest average summer temperature. On the contrary, higher air temperatures from February to mid-April are the cause of the vegetation season early beginning and are related to the increment increase. Castaldi et al. (2019) mention that the minimum temperature in February and March plays a key role for Douglas-fir, too. High precipitation sum at the beginning of the vegetation season and in July is the most significant for Douglas-fir. Similar conclusion was achieved by Vejpustková and Čihák (2019): while Norway spruce was negatively affected by temperatures in the summer months, Douglas-fir growth was positively correlated with temperatures in February and March. Generally, June and July are the most important months in terms of climate effect on radial growth and xylem formation of the studied tree species (Little et al., 2008; Putalová et al., 2019).
The air humidity has also a great effect on both tree species. This indicates sufficient precipitation and their even distribution. The soil attributes, local climatic characteristics and especially air humidity are necessary to be taken into account in connection with the increase of hot and dry periods, when selecting localities to Norway spruce substitution by Douglas-fir. Radial growth is also affected by the intensity of dry periods, both in Norway spruce and Douglas-fir, as published in a study by Sergent et al. (2014). Also, according to Littel et al. (2008), an increase in temperature in April to September without an increase in summer precipitation or soil moisture reserves causes probable Douglas-fir growth decrease.
Many stands in the Písek region, where Norway spruce is the dominant tree species, are falling apart at present. It is very weak, ceases to be resistant to spruce bark beetle attacks and it is slowly disappearing from the forests in this region due to extreme droughts during previous years. Similarly, rapid decline of Norway spruce was observed in recent years across Europe (Grodzski et al., 2010; Hlásný and Sitková, 2010; Vacek et al., 2019b; Toth et al., 2020). Extensive spruce disturbances are caused by the increasing frequency of extreme climatic events (long-term droughts, windstorms, etc.) during climate change (Schelhaas et al., 2003; Krejčí et al., 2013; Prokůpková et al., 2020; Šimůnek et al., 2020). On the other hand, Douglas-fir appears to be coping with this situation and remains more or less vital in stands, while experiencing a small increment reduction. Study from Germany also showed that Douglas-fir is a more tolerant tree species to extreme droughts compared to Norway spruce (Vitali et al., 2018). Moreover, spruce stands have shown historically low resistance and high damage as a result of air pollution load (Vacek et al., 2015, 2017; Králíček et al., 2017).
According to analysis of negative pointer year with extremely low radial growth, Norway spruce was also significantly less resistant to climate extremes compared to Douglas-fir (4 vs. 7 years). Positive years for both tree species were then 1997 and 2002 and negative years 1976 and 2018. Kern et al. (2017) reported 2000 as a year with extreme negative anomalies for coniferous in the Czech Republic and subsequently 2001, as in our case for Norway spruce. On the other hand, this work states, as the most positive anomaly year 2014, when this year was very good for spruce but not as much for Douglas-fir in our case. Furthermore, negative year 1993 was found for Norway spruce, similarly to the Jeseníky Mountains (Mikulenka et al., 2020). Another agreement with other works is the negative year 2015, which was characterized by extremely high temperatures with low amounts of precipitation in the summer months not only in our case, but also in the Giant Mountains (Vacek et al., 2020c). Tree growth in terms of both positive and negative anomalies shows strong land cover and soil dependency (Kern et al., 2017; Šimůnek et al., 2021). Because both species show different reaction to climatic extremes, it is possible that a mixture stand would be more resistant in terms of growth stability than monospecific stands. This is also confirmed by the different significant pointer years affecting radial growth, where out of a total of 20 years, only 3 years were the same for both tree species. The advantage of mixed forests in terms of stability and resilience to climate change and droughts was also confirmed by other studies (Vacek et al., 2019c, 2020a; Hájek et al., 2020; Pretzsch et al., 2020). Moreover, the highest productivity potential was observed in species-rich forest stands in our study (higher basal area by 13.1%). Pretzsch and Thurm (2016) found that mixtures of Douglas-fir and European beech showed overyielding in comparison to those species in pure stands. The results also suggest that research focused on potential use of Douglas-fir in single-selection silvicultural system in Central Europe could be interesting, similarly to Scots pine in Spain (Gallo et al., 2020b).
Presented correlations cannot be interchanged with causality (Mason 2004). Causality is the subject of a possible explanation of these connections, it cannot be directly derived from the data. Substantial finding in our study is that the strongest effect can be expected in the average humidity for both tree species, and only then for the total precipitation; average temperatures have a weaker effect. Because the correlations are based on a large amount of data, even (in absolute value) small coefficients r are statistically significant (see note to Table 3; all values with | r | > 0.004 are significant at level α = 0.05). Low degree of explanation of data variance (x2) is related to variability between individuals, between areas, localities, with the influence of other factors and finally with the measurement error (climate was not measured on research plots but on the stations): from this point of view up to approximately 9% of the explained variability for Douglas-fir (average humidity) and 10% for Norway spruce is not so low. In addition, the effect of the combination of maximum and minimum correlation in the form (rmin2 + rmax2) can be considered, reaching up to 12% for Douglas-fir (average humidity).
5. Conclusion
Both Pseudotsuga menziesii and Picea abies in the region show a strong influence of the locality. It is valid especially for Picea abies, which indicates that it grows in unsuitable conditions here and its existence in these altitudes is strongly endangered. On the other hand, Douglas-fir is more resistant to climate extremes and also confirmed a higher productivity potential. However, both species are potentially influenced by climate in different manner. The correlation of average humidity can be considered strongest for both tree species, second most significant results were for the total precipitation, average temperatures showed a weaker significance. High temperatures together with lack of precipitation in the first part of the growing period proved to have a negative effect on the growth of both Norway spruce and Douglas-fir. Spruce coped better with low temperatures and was more dependent on the course of precipitation during the year. Further research of cultivating Douglas-fir and Norway spruce, particularly in coniferous mixtures, is needed to evaluate different ways of adapting the forest stands to various climatic extremes and ongoing climate change.
Acknowledgements
The study was supported by the Ministry of Agriculture of the Czech Republic (project No. QK1920328). The paper was elaborated using the facilities at Truba Research Station in Kostelec nad Černými lesy (Department of Silviculture, Faculty of Forestry and Wood Science, Czech University of Life Sciences Prague).
References
Acosta-Hernández A.C., Pompa-García M., Camarero J.J. 2017. An Updated Review of Dendrochronological Investigations in Mexico, a Megadiverse Country with a High Potential for Tree-Ring Sciences. Forests 2017(8): 160.
Arreola-Ortiz, M.R.; Návar-Cháidez, J.D.J. 2010. Análisis de sequías y productividad con cronologías de Pseudotsuga menziesii Rob. & Fern., y su asociación con El Niño en el nordeste de México. Investigaciones geográficas 71: 7–20.
Baláš M., Kuneš I., Gallo J., Rašáková N. 2016. Review on Betula oycoviensis and foliar morphometry of the species in Volyně, Czech Republic. Dendrobiology 76: 117–125.
Białobok S, Mejnartowicz L 1970. Provenance differentiation among Douglas fir seedlings. Arboretum Kórnickie 15: 197–219.
Brundu G., Pauchard A., Pyšek P., Pergl J., Bindewald A.M., Brunori A., Canavan S., Campagnaro T., Celesti-Grapow L., de Sá Dechoum M., Dufour-Dror J.M., Essl F., Flory S. L., Genovesi P., Guarino F., Guangzhe L., Hulme P., Jäger H., Kettle J. C., Krumm F., Langdon B., Lapin K., Lozano V., Le Roux J. J., Novoa A., Nuñez M. A., Porté A. J., Silva J. S., Schaffner U., Sitzia T., Tanner R., Tshidada N., Vítková M., Westergren M., Wilson J. R. U., Richardson D. 2020. Global guidelines for the sustainable use of non-native trees toprevent tree invasions and mitigate their negative impacts. NeoBiota 61: 65–116.
Brus R., Pötzelsberger E., Lapin K., Brundu G., Orazio C., Straigyte L., Hasenauer H. 2019. Extent, distribution and origin of non-native forest tree species in Europe. Scandinavian Journal of Forest Research 34(7): 533–544.
Carrillo-Gavilán A., Espelta J. M., Vilà M. 2012. Establishment constraints of an alien and a native conifer in different habitats. Biological Invasions 14(6): 1279–1289.
Castaldi C., Vacchiano G., Marchi M., Corona P., 2017. Projecting Nonnative Douglas Fir Plantations in Southern Europe with the Forest Vegetation Simulator. Forest Science 63(1):101–110.
Castaldi C., Marchi M., Vacchiano G., Corona P. 2019. Douglas-fir climate sensitivity at two contrasting sites along the southern limit of the European planting range. Journal of Forestry Research 31(6): 2193–2204.
Čater M. 2021. Microsites Influence the Light Response of Young Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco). Forests 12(6): 687.
Chakraborty D., Matulla Ch., Andre K., Weissenbacher L., Schueler S. 2019. Survival of Douglas-fir provenances in Austria: site-specific late and early frost events are more important than provenance origin. Annals of Forest Science 76 (4): 100.
Chylarecki H. 1976. Badania nad daglezja w Polsce w różnych warunkach ekologicznych (Research on Douglas fir in Poland under different ecological conditions). Arboretum Kórnickie 21: 15–123 (in Polish).
Cukor J., Baláš M., Kupka I., Tužinský M. 2017a. The condition of forest stands on afforested agricultural land in the Orlické hory Mts. Journal of Forest Science 63(1): 1–8.
Cukor J., Linhart L., Vacek Z., Baláš M., Linda R. 2017b. The effects of Alginite fertilization on selected tree species seedlings performance on afforested agricultural lands. Central European Forestry Journal 63(1): 48–56.
Cukor J., Zeidler A., Vacek Z., Vacek S., Šimůnek V., Gallo J. 2020. Comparison of growth and wood quality of Norway spruce and European larch: effect of previous land use. European Journal of Forest Research 139(3): 1–14.
Desplanque C., Rolland C., Schweingruber F.H. 1999. Influence of species and abiotic factors on extreme tree ring modulation. Trees 13(4): 218–227.
Eberhard B., Hasenauer H. 2018. Modeling Regeneration of Douglas fir forests in Central Europe. Austrian Journal of Forest Science 135(1): 33–51.
Eckhart T., Walcher S., Hasenauer H., Van Loo M. 2017. Genetic diversity and adaptive traits of European versus American Douglas-fir seedlings. European Journal of Forest Research 136(5): 811–825.
Eckhart T., Pötzelsberger E., Koeck R., Thom D., Lair G.J., van Loo M., Hasenauer H. 2019. Forest stand productivity derived from site conditions: an assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Annals of Forest Science 76(1): 19.
Essl F. 2005. Distribution, status and habitat preference of subspontaneous Douglas fir stands (Pseudotsuga menziesii) in Austria (Germ.). Phyton-annales Rei Botanicae 45(1): 117–143.
Feliksik E., Wilczyński S. 2004. Dendroclimatic regions of Douglas fir Pseudotsuga menziesii (Mirb.) Franco in western and northern Poland. Dendrobiology 52: 9–15.
Fischer S., Neuwirth B. 2012. Climate sensitivity of Douglas-fir in Eifel and Kellerwald. Allgemeine Forst und Jagdzeitung 183(1–2): 23–33.
Gallo J., Kuneš I., Baláš M., Nováková O., Drury M. L. 2014. Occurrence of frost episodes and their dynamics in height gradient above the ground in the Jizerské hory Mts. Journal of Forest Science 60(1): 35–41.
Gallo J., Baláš M., Linda R., Kuneš I. 2017. Growth performance and resistance to near-ground late frosts of Fagus sylvatica L. plantation treated by a brassinosteroid compound. Journal of Forest Science 63(3): 117–125.
Gallo J., Baláš M., Linda R., Kuneš I. 2020a. The effects of planting stock size and weeding on survival and growth of small-leaved lime under drought-heat stress in the Czech Republic. Austrian Journal of Forest Science 137(1): 43–66.
Gallo J., Bílek L., Šimůnek V., Roig S., Fernández J. A. B. 2020b. Uneven-aged silviculture of Scots pine in Bohemia and Central Spain: comparison study of stand reaction to transition and long-term selection management. Journal of Forest Science 66(1): 22–35.
George J-P., Grabner M., Campelo F., Karanitsch-Ackert S., Kayer K., Klumpp R.T., Schüler S. 2019. Intra-specific variation in growth and wood density traits under water-limited conditions: Long-term-, short-term-, and sudden responses of four conifer tree species. Science of Total Environment 660: 631–643.
Giorgi F., Coppola E. 2007. European climate-change oscillation (ECO). Geophysical Research Letters 34 (21): [art. no. L21703].
Gömöry D., Krajmerová D., Hrivnák M., Longauer R. 2020. Assisted migration vs. close-to-nature forestry: what are the prospects for tree populations under climate change? Central European Forestry Journal 66: 63–70.
Grodzki W. 2010. The decline of Norway spruce Picea abies (L.) Karst. stands in Beskid Śląski and Żywiecki: Theoretical concept and reality. Beskydy 3: 19–26.
Grissino-Mayer H. D., Holmes R. L., Fritts H. C. 1992. International tree-ring data bank program library: user´s manual. Laboratory of Tree-Ring Research, University of Arizona, Tuscon, USA.
Hájek V., Vacek Z., Vacek S., Bílek L., Prausová R., Linda R., Bulušek D., Králíček I. 2020. Changes in diversity of protected scree and herb-rich beech forest ecosystems over 55 years. Central European Forestry Journal 66(4): 202–217.
Harlow W. M., Harrar E. S., White F. M., 1979. Textbook of dendrology. Covering the Important Forest Trees of the United States and Canada. Ed. 6. McGraw-Hill Book Company: 140–146.
Hedlund K., Regina I. S., Van der Putten W.H., Lepš J., Díaz T., Korthals G.W., Lavorel S., Brown V.K., Gormsen D., Mortimer S.R., Barrueco C.R., Roy J., Smilauer P., Smilauerová M., Van Dijk C. 2003. Plant species diversity, plant biomass and responses of the soil community on abandoned land across Europe: idiosyncracy or above-belowground time lags. Oikos 103: 45–58.
Hermann R. K., Lavender D. P. 1999. Douglas-fir planted forests. New Forests 17(1–3): 53–70.
Hintsteiner W. J., van Loo M., Neophytou C., Schueler S., Hasenauer H. 2018. The geographic origin of old Douglas-fir stands growing in Central Europe. European Journal of Forest Research 137(4): 447–461.
Hlásny T., Sitková Z. 2010. Spruce forests decline in the Beskids. National Forest Centre – Forest Research Institute Zvolen & Czech University of Life Sciences Prague & Forestry and Game Management Research Institute Jíloviště – Strnady, Zvolen, 182 p.
Hofman J. 1964. Pěstování douglasky. SZN, Praha, 253 p.
Kacálek D., Novák J., Bartoš J., Slodičák M., Balcar V., Černohous V. 2010. Vlastnosti nadložního humusu a svrchní vrstvy půdy ve vztahu k druhům dřevin. Zprávy lesnického výzkumu 55(1): 19–25.
Kern A., Marjanović H., Dobor L., Anić M., Hlásny T., Barcza Z. 2017. Identification of years with extreme vegetation state in Central Europe based on remote sensing and meteorological data. South-east European forestry: SEEFOR 8(1): 1–20.
Knibbe B. 2007. PAST4: personal analysis system for treering research, Version 4.2. SCIEM, Vienna.
Körner Ch. 2005. An introduction to the functional diversity of temperate forest trees. In: Scherer-Lorenzen, Körner Ch., Schulze E.-D. (eds.) Forest diversity and function. Temperate and boreal systems. Ecological Studies 176: 13–37. Springer-Verlag, Berlin et Heidelberg, 389 p.
Köppen W. 1936. Das Geographische System der Klimate, Handbuch der Klimatologie. Gebrüder Borntraeger, Berlin.
Kraft G. 1884. Beiträge zur zur Lehre von den Durchforstungen. Schlagstellungen und Lichtungshieben. Klindworth, Hannover.
Král J., Vacek S., Vacek Z., Putalová T., Bulušek D., Štefančík I. 2015. Structure, development and health status of spruce forests affected by air pollution in the western Krkonoše Mts. in 1979–2014. Central European Forestry Journal 61(3): 175–187.
Králíček I., Vacek Z., Vacek S., Remeš J., Bulušek D., Král J., Štefančík I., Putalová T. 2017. Dynamics and structure of mountain autochthonous spruce-beech forests: impact of hilltop phenomenon, air pollutants and climate. Dendrobiology 77: 119–137.
Krejčí F., Vacek S., Bílek L., Mikeska M., Hejcmanová P., Vacek Z. 2013. The effects of climatic conditions and forest site types on disintegration rates in Picea abies occurring at the Modrava Peat Bogs in the Šumava National Park. Dendrobiology 70:35–44.
Kšír J., Beran F., Podrázský V., Novotný P., Dostál J., Kubeček J. 2015. Results of the evaluation of the provenance research plot with Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) on the locality Hůrky (Southern Bohemia) at the age of 44 years. Zprávy lesnického výzkumu 60(2): 104–114.
Kubeček J., Štefančík I., Podrázský V., Longauer R. 2014. Results of the research of Douglas-fir in the Czech Republic and Slovakia: a review. Central European Forestry Journal 60(2): 116–124.
Kuneš I., Baláš M., Linda R., Gallo J., Nováková O. 2017. Effects of brassinosteroid application on seed germination of Norway spruce, Scots pine, Douglas fir and English oak. iForest-Biogeosciences and Forestry 10(1): 121.
Kuneš I., Baláš M., Gallo J., Šulitka M., Pinidiya Arachchilage C. 2019. Trnovník akát (Robinia pseudoacacia) a jeho role ve středoevropském a českém prostoru: review. Zprávy lesnického výzkumu, 64(4): 181–190.
Kupka I., Podrázský V. 2010. Vliv druhového složení porostů na zalesněné zemědělské půdě na pedofyzikální vlastnosti a poutání uhlíku v povrchových horizontech [Effects of species composition of forest stands on afforested agricultural land on the soil physical properties and C-fixation in the surface horizons]. In: Knott R., Peňáz J., Vaněk P. (eds.) Silviculture at lower forest vegetation zones: 71–76. Mendelova univerzita v Brně, 172 p.
Kupka I., Podrázský V., Kubeček J. 2013. Soil-forming effect of Douglas fir at lower altitudes. Journal of Forest Science 59(9): 345–351.
Lachenbruch B., Johnson R. G. 2020. Different radial growth rate effects on outerwood properties of coastal Douglas-fir in healthy trees vs. trees impacted by Swiss Needle Cast. Canadian Journal of Forest Research (first online).
Lepš J. 2004. Variability in population and community biomass in a grassland community affected by environmental productivity and diversity. Oikos 107: 64–71.
Littell J., Peterson D. L., Tjoelker M. 2008. Douglas-fir growth in mountain ecosystems: water limits tree growth from stand to region. Ecological Monographs 78(3): 349–368.
Macků J., Kosová D. 2020. Scenario of the climate change model HadGem in the regional forest development plans. Zprávy lesnického výzkumu 65(1): 28–39.
Mäkinen H., Nöjd P., Kahle H. P., Neumann U., Tveite B., Mielikäinen K., Röhle H., Spiecker H. 2002. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. Forest Ecology and Management 171(3): 243–259.
Mason E. 2012. Designing silvicultural regimes with a structural log index. New Zealand Journal of Forestry 57(2): 13–8.
Matějka K., Podrázský V., Viewegh J., Martiník A. 2015. Srovnání bylinné etáže v porostech douglasky tisolisté (Pseudotsuga menziesii (Mirb.) Franco) a v porostech jiných dřevin. Zprávy lesnického výzkumu 60(3): 201–210.
Matějka K. 2020. Nápověda k programu DBreleve. Databáze fytocenologických snímků, verze 2.5. – URL: https://www.infodatasys.cz/software/hlp_dbreleve/dbreleve.htm
Mauer O., Palátová E. 2012. Root system development in Douglas-fir (Pseudotsuga menziesii /Mirb./ Franco) on fertile sites. Journal of Forest Science 58(9): 400–409.
Mauer O., Vaněk P. 2014. Douglaska tisolistá (Pseudotsuga menziesii /Mirb./ Franco) jako meliorační a zpevňující dřevina, růst douglasky na různých stanovištích od 2. do 7. lesního vegetačního stupně [Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) as amelioration and reinforcement tree species, Douglas-fir growth at various habitats from 2nd to 7th forest vegetation zones]. In: Slabý R. (ed.) Douglas-fir – Tree of the year 2014: 18-34. Česká lesnická společnost o.s., Praha, 204 p. (available in English at https://www.infodatasys.cz/proj009/Douglasfir2014tr.pdf)
Mejnartowicz L. 1976. Genetic investigations on Douglas–fir (Pseudotsuga menziesii (Mirb.) Franco) populations. Arboretum Kórnickie 21: 126-179.
Mlíkovský J., Stýblo P., eds., 2006. Nepůvodní druhy fauny a flóry ČR, ČSOP Praha, 496 pp.
Menšík L., Kulhavý J., Kantor P., Remeš M. 2009. Humus conditions of stands with the different proportion of Douglas-fir in training forest district Hůrky and the Křtiny Forest Training Enterprise. Journal of Forest Science 55(8): 345–356.
Mikulenka P., Prokůpková A., Vacek Z., Vacek S., Bulušek D., Simon J., Šimůnek V., Hájek V. 2020. Effect of climate and air pollution on radial growth of mixed forests: Abies alba Mill. vs. Picea abies (L.) Karst. Central European Forestry Journal 66(1): 23–36.
Mondek J., Baláš M. 2019. Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) and its role in the Czech forests. Journal of Forest Science 65(2): 41–50.
MZE 2013. Zpráva o stavu lesa a lesního hospodářství v roce 2012. MZE, Praha, 132 p.
MZE 2020. Zpráva o stavu lesa a lesního hospodářství v roce 2019. MZE, Praha, 124 p.
Nadezhdina N., Urban J., Čermák J., Nadezhdin V., Kantor P. 2014. Comparative study of long-term water uptake of Norway spruce and Douglas-fir in Moravian upland. Journal of Hydrology and Hydromechanics 62: 1–6.
Němeček J., Macků J., Vokoun J., Vavříček D., Novák P. 2001. Taxonomický klasifikační systém půd České republiky [The Czech taxonomic soil classification system]. Česká zamědělská univerzita v Praze, 79 p.
Neophytou C., van Loo M., Hasenauer H. 2020. Genetic diversity in introduced Douglas-fir and its natural regeneration in Central Europe. Forestry: An International Journal of Forest Research 93(4): 535–544.
Novák J., Kacálek D., Podrázský V., Šimerda L. (eds.) 2018. Uplatnění douglasky tisolisté v lesním hospodářství ČR. Lesnická práce s.r.o., Kostelec nad Černými lesy, 216 p.
Novák J., Dušek D., Kacálek D. 2019: Growth of juvenile Douglas-fir mixed with native tree species on different forest sites. Zprávy lesnického výzkumu 64(3): 133–139.
Palátová P., Dudík R., Hušbauer, J. 2017. Czech wood and potential of its consumption in the Czech republic. In: Dudík R. (ed.) More wood, better management, increasing effectiveness: Starting points and perspective: 33–37. Czech University of Life Sciences Prague, 245 p.
Pergl et al. 2016: Black, Grey and Watch Lists of alien species in the Czech Republic based on environmental impacts and management strategy. NeoBiota 28: 1–37.
Petkova K., Georgieva M., Uzunov M. 2014. Investigation of Douglas-fir provenance test in North-Western Bulgaria at the age of 24 years. Journal of Forest Science 60(7): 288–296.
Perić S., Tijardović M., Jazbec A. 2011. Results of research on douglas fir provenances in ecologicaly different continenetal parts of Croatia. Šumarski List 135(13): 190–201.
Podrázský V., Remeš J. 2008. Půdotvorná role významných introdukovaných jehličnanů – douglasky tisolisté, jedle obrovské a vejmutovky. Zprávy lesnického výzkumu 53(1): 27–34.
Podrázský V., Kupka I. 2011. Vliv douglasky tisolisté (Psedotsuga menziesii /Mirb./ Franco) na základní pedofyzikální charakteristiky lesních půd. Zprávy lesnického výzkumu 56 (Special): 1–5.
Podrázský V., Čermák R., Zahradník D., Kouba J. 2013. Production of Douglas-fir in the Czech Republic based on national forest inventory data. Journal of Forest Science 59(10): 398–404.
Podrázský V., Zahradník D., Remeš J. 2014a. Potential consequences of tree species and age structure changes of forests in the Czech republic – Review of forest inventory data. Wood Research 59(3): 483–490.
Podrázský V., Martiník A., Matějka K., Viewegh J. 2014b. Effects of Douglas-fir (Pseudotsuga menziesii [Mirb.] Franco) on understorey layer species diversity in managed forests. Journal of Forest Science 60(7): 263–271.
Podrázský V., Noha M., Kubeček J. 2015. Contribution for determination of the optimal share of Douglas-fir in the forest stands. In: Houšková K., Černý J. (eds.) Proceedings of Central European Silviculture: 208–217. Mendelova univerzita v Brně, 222 p.
Podrázský V., Remeš J., Sloup R., Pulkrab K., Novotný S. 2016. Douglas-fir – Partial substitution for declining conifer timber supply – Review of Czech data. Wood Research 61(4): 525–530.
Podrázský V. 2016. Potential substitution of Norway spruce by the Douglas-fir in the Czech Republic. In: Kacálek D., Novák J., Nováková K., Součková J. (eds.): Forest functions in Changing Environments: 99-104. Studijní středisko Ústavu jazykové a odborné přípravy Univerzity Karlovy, Praha, 246 p.
Podrázský V., Vacek Z., Kupka I., Vacek S., Třeštík M., Cukor J. 2018. Effects of silver fir (Abies alba Mill.) on the humus forms in Norway spruce (Picea abies (L.) H. Karst.) stands. Journal of Forest Science 64(6): 245–250.
Podrázský V., Kupka I., Prknová H. 2020. Substitution of Norway spruce for Douglas-fir: changes of soil microbial activities as climate change induced shift in species composition, a case study. Central European Forestry Journal 66(2): 71–77.
Poleno Z., Vacek S., Podrázský V., Remeš J., Štefančík I., Mikeska M., Kobliha J., Kupka I., Malík V., Turčáni M., Dvořák J., Zatloukal V., Bílek L., Baláš M., Simon J. 2009. Pěstování lesů III. Praktické postupy pěstování lesů. [Silviculture III. Practical methods in silviculture]. Kostelec nad Černými lesy, Lesnická práce, s.r.o., 952 p.
Popov E. B. 2014. Results of 20 years old Douglas-fir provenance experiment established on the northern slopes of Rila Mountain in Bulgaria. Journal of Forest Science 60(9): 394–399.
Pötzelsberger E., Eckhart T., Hasenauer H. 2019. Wachstumspotenziale für Douglasie in Österreich und Deutschland. [Mapping the growth potential of Douglas-fir in Austria and Germany]. Austrian Journal Forest Science 136(2): 69–86.
Pötzelsberger E., Spiecker H., Neophytou C., Mohren F., Gazda A., Hasenauer H. 2020. Growing non-native trees in European forests brings benefits and opportunities but also has its risks and limits. Current Forestry Reports 6(4): 339–353.
Pretzsch H., Grams T., Häberle K. H., Pritsch K., Bauerle T., Rötzer T. 2020. Growth and mortality of Norway spruce and European beech in monospecific and mixed-species stands under natural episodic and experimentally extended drought. Results of the KROOF throughfall exclusion experiment. Trees 34(4): 957–970.
Prokůpková A., Vacek Z., Vacek S., Blažejová J., Schwarz O., Bulušek D. 2020. Dynamics of natural regeneration of mountain forests after wind disturbance: model study for the Krkonoše Mts. (Czech Republic). Zprávy Lesnického Výzkumu 65(2): 72–81.
Pulkrab K., Sloup R., Podrázský V. 2015. Production potential of the forests in the Czech republic. BioResources 10(3): 4711–4725.
Putalová T., Vacek Z., Vacek S., Štefančík I., Bulušek D., Král J. 2019. Tree-ring widths as an indicator of air pollution stress and climate conditions in different Norway spruce forest stands in the Krkonoše Mts. Central European Forestry Journal 65(1): 21–33.
Pyšek P., Chytrý M., Pergl J., Sádlo J., Wild J. 2012. Plant invasions in the Czech Republic: current state, introduction dynamics, invasive species and invaded habitats. Preslia 84: 576–630.
Remeš J., Bílek L., Novák J., Vacek Z., Vacek S., Putalová T., Koubek L. 2015. Diameter increment of beech in relation to social position of trees, climate characteristics and thinning intensity. Journal of Forest Science 61: 456–464.
Remeš J., Pulkrab K., Bílek L., Podrázský V. 2020. Economic and production effect of tree species change as a result of adaptation to climate change. Forests 11(4): 431.
Riedl M., Jarský V., Palátová P., Sloup R. 2019. The Challenges of the Forestry Sector Communication Based on an Analysis of Research Studies in the Czech Republic. Forests 10(11): 935.
Sageman B.B., Hollander D.H. 1999. Cross correlation of paleoecological and geochemical proxies: A holistic approach to the study of past global change. In: E. Barrera and C.C. Johnson (eds.), GSA Special Paper 332, pp. 365–384.
Schär C., Vidale P. L., Lüthi D., Frei C., Häberli C., Liniger M. A., Appenzeller C. 2004. The role of increasing temperature variability in European summer heatwaves. Nature 427(6972): 332–336.
Schmid M., Pautasso M., Holdenrieder O. 2014. Ecological consequences of Douglas fir (Pseudotsuga menziesii) cultivation in Europe. European Journal of Forest Research 133(1): 13–29.
Schweingruber F.H. 1996. Tree Rings and Environment Dendroecology. Swiss Federal Institute for Forest, Snow and Landscape Research, Birmensdorf, Switzerland, 609 p.
Schelhaas M.J., Nabuurs G.J., Schuck A. 2003. Natural disturbances in the European forests in the 19th and 20th centuries. Global Change Biology 9(11): 1620–1633.
Schober R. 1963. Erfahrungen mit der Douglasie in Europa. Allgemeine Forstzeitschrift 18(30): 473–519 (in German).
Sergent A. S., Rozenberg P., Marcais B., Lefevre Y., Bastien J. C., Nageleisen, L. M. et al. 2010. Vulnerability of Douglas-fir in a changing climate: study of decline in France after the extreme 2003´s drought. In: Spiecker H., Kohnle U., Makkonen-Spiecker K., Brill C. (eds.) Opportunities and risks for Douglas fir in a changing climate: 21–22. Eigenverlag der FVA, Freiburg, Berichte Frei¬burger Forstliche Forschung (85).
Sergent A. S., Rozenberg P., Breda N. 2014. Douglas-fir is vulnerable to exceptional and recurrent drought episodes and recovers less well on less fertile sites. Annals of Forest Science 71(6): 697–708.
Slodičák M., Novák J., Mauer O., Podrázský V. (eds.) 2014. Pěstební postupy při zavádění douglasky do porostních směsí v podmínkách ČR. Lesnická práce s.r.o., Kostelec nad Černými lesy, 272 p.
Šach F., Černohous V., Kacálek D. 2019. Water regime of Douglas-fir and forest stands with Douglas-fir: review. Zprávy lesnického výzkumu 64(3): 149–154.
Šimůnek V., Vacek Z., Vacek S. 2020. Solar Cycles in Salvage Logging: National Data from the Czech Republic Confirm Significant Correlation. Forests 11(9): 973.
Šimůnek V., Vacek Z., Vacek S., Ripullone F., Hájek V., D’Andrea G. 2021. Tree Rings of European Beech (Fagus Sylvatica L.) Indicate the Relationship with Solar Cycles During Climate Change in Central and Southern Europe. Forests 12(3): 259.
Šindelář J. 2003. Aktuální problémy a možnosti pěstování douglasky tisolisté. Lesnická práce 82(5): 14–16.
Špačková I., Lepš J. 2001. Procedure for separating the selection effect from other effects in diversity-productivity relationship. Ecology Letters 4(6): 585–594.
Štícha V., Sharma R. P., Vacek Z., Vacek S., Nuhlíček O. 2019. Timber and branch volume prediction: Effects of stand and site characteristics on dendromass and timber-to-branch volume ratio of Norway spruce in managed forests. Forests 10(2): 144.
Thurm E. A., Pretzsch H., 2016. Improved productivity and modified tree morphology of mixed versus pure stands of European beech (Fagus sylvatica) and Douglas-fir (Pseudotsuga menziesii) with increasing precipitation and age. Annals of Forest Science 73(4): 1047–1061.
Tschopp T., Holderegger R., Bollmann K. 2015. Auswirkungen der Douglasie auf die Waldbiodiversität. Schweizerische Zeitschrift fur Forstwesen 166(1): 9–15.
Toth D., Maitah M., Maitah K., Jarolínová V. 2020. The Impacts of Calamity Logging on the Development of Spruce Wood Prices in Czech Forestry. Forests 11(3): 283.
Ulbrichová I., Kupka I., Podrázský V., Kubeček J., Fulín M. 2014. Douglaska jako meliorační a zpevňující dřevina. Zprávy lesnického výzkumu 59(1): 72–78.
Urban J., Čermák J., Kantor P. 2011. Srovnání radiálního růstu a transpirace douglasky tisolisté a smrku ztepilého. In: Prknová H. (ed.) Aktuality v pěstování méně častých dřevin v České republice: 77–81. Česká zemědělská univerzita v Praze, 118 p.
Vacek S., Hůnová I., Vacek Z., Hejcmanová P., Podrázský V., Král J., Putalová T., Moser, W. K. 2015. Effects of air pollution and climatic factors on Norway spruce forests in the Orlické hory Mts.(Czech Republic), 1979–2014. European Journal of Forest Research 134(6): 1127–1142.
Vacek S., Černý T., Vacek Z., Podrázský V., Mikeska M., Králíček I. 2017. Long-term changes in vegetation and site conditions in beech and spruce forests of lower mountain ranges of Central Europe. Forest Ecology and Management 398: 75–90.
Vacek S., Prokůpková A., Vacek Z., Bulušek D., Šimůnek V., Králíček I., Prausová R., Hájek, V. 2019a. Growth response of mixed beech forests to climate change, various management and game pressure in Central Europe. Journal of Forest Science 65(9): 331–345.
Vacek S., Vacek Z., Ulbrichová I., Remeš J., Podrázský V., Vach M., Bulušek D., Král J., Putalová, T. 2019b. The effects of fertilization on the health status, nutrition and growth of Norway spruce forests with yellowing symptoms. Scandinavian Journal of Forest Research 34(4): 267–281.
Vacek S., Vacek Z., Ulbrichová I., Bulušek D., Prokůpková A., Král J., Vančura K. 2019c. Biodiversity dynamics of differently managed lowland forests left to spontaneous development in Central Europe. Austrian Journal of Forest Science 136(3): 249–281.
Vacek Z., Vacek S., Bílek L., Král J., Remeš J., Bulušek D., Králíček I. 2014. Ungulate impact on natural regeneration in spruce-beech-fir stands in Černý důl Nature Reserve in the Orlické Hory Mountains, case study from Central Sudetes. Forests 5(11): 2929–2946.
Vacek Z. 2017. Structure and dynamics of spruce-beech-fir forests in Nature Reserves of the Orlické hory Mts. in relation to ungulate game. Central European Forestry Journal 63(1): 23–34.
Vacek Z., Vacek S., Slanař J., Bílek L., Bulušek D., Štefančík I., Králíček I., Vančura, K. 2019c. Adaption of Norway spruce and European beech forests under climate change: from resistance to close-to-nature silviculture. Central European Forestry Journal 65(2): 129–144.
Vacek Z., Cukor J., Linda R., Vacek S., Šimůnek V., Brichta J., Gallo J., Prokůpková A. 2020c. Bark stripping, the crucial factor affecting stem rot development and timber production of Norway spruce forests in Central Europe. Forest Ecology and Management 474: 118360.
Vacek Z., Prokůpková A., Vacek S., Cukor J., Bílek L., Gallo J., Bulušek D. 2020a. Silviculture as a tool to support stability and diversity of forests under climate change: study from Krkonoše Mountains. Central European Forestry Journal 66(2): 116–129.
Vacek Z., Vacek S., Prokůpková A., Bulušek D., Podrázský V., Hůnová I., Putalová T., Král J. 2020b. Long-term effect of climate and air pollution on health status and growth of Picea abies (L.) Karst. peaty forests in the Black Triangle region. Dendrobiology 83: 1–19.
Vacek Z., Prokůková A., Vacek S., Bulušek D., Šimůnek V., Hájek V., Králíček I. 2021. Effect of Norway spruce and European beech mixing in relation to climate change: structural and growth perspectives of mountain forests in Central Europe. Forest Ecology and Management (in press).
van Loo M., Lazic D., Chakraborty D., Hasenauer H., Schüler S. 2019. North American Douglas-fir (P. menziesii) in Europe: establishment and reproduction within new geographic space without consequences for its genetic diversity. Biological invasions 21(11): 3249–3267.
Vašíček J. 2014. Douglaska tisolistá v číslech [Douglas-fir in numbers]. In: Slabý R. (ed.) Douglas-fir – Tree of the year 2014: 5-10. Česká lesnická společnost o.s., Praha, 204 p. (available in English at www.infodatasys.cz/proj009/Douglasfir2014tr.pdf)
Vejputková M., Čihák T. 2019. Climate response of douglas fir reveals recently increased sensitivity to drought stress in Central Europe. Forests 10(2): 97.
Viewegh J., Kusbach A., Mikeska M. 2003. Czech forest ecosystem classification. Journal of Forest Science 49(2): 74–82.
Viewegh J., Podrázský V., Matějka K. 2014. Charakterystyka roslinnosci runa ksztaltujacej sie pod drzewostanami daglezjovymi (Pseudotsuga menziesii (Mirb.) Franco) w lasach gospodarczych Republiki Czeskiej. Sylwan 158(4): 277–284.
Vitali V., Büntgen U., Bauhus J. 2017. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Global Change Biology 23(12): 5108–5119.
Vitali V., Forrester D. I., Bauhus J. 2018. Know your neighbours: drought response of Norway Spruce, Silver Fir and Douglas Fir in mixed forests depends on species identity and diversity of tree neighbourhoods. Ecosystems 21(6): 1215–1229.
Wilczyński S., Feliksik E. 2007: Local chronologies and regional diversity of dendrochronological signal od douglas fir in Poland. Geochronometria 26: 69–80.
Yamaguchi, D.K. 1991. A simple method for cross-dating increment cores from living trees. Canadian Journal Forest Research 21(3): 414–416.
Zeller B., Andrianarisoa S., Jussy J. H. 2010. Impact of Douglas-fir on the N cycle: Douglas fir promote nitrification? In: Spiecker H., Kohnle U., Makkonen-Spiecker K., Brill C. (eds.) Opportunities and risks for Douglas fir in a changing climate: 11. Eigenverlag der FVA, Freiburg, Berichte Freiburger Forstliche Forschung 85.



