Schlüsselbegriffe: Wasserpotential, Prolin, Chlorophyll, Traubeneiche, Kohlenhydrate, Türkei
Available at https://doi.org/10.53203/fs.2402.1
See below the issue 2/2024 as E-Paper or have a look at our E-Paper archive dating back to 1955.
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Abstract
Due to climate change, forest tree species are expected to be impacted by drought. As drought becomes a worldwide problem, there is a need to investigate the responses of forest tree species to drought. In this study, the objectives were
(I) to determine the physiological and biochemical responses of seedlings grown from seeds obtained from different populations of Quercus petraea subsp. iberica naturally distributed in Turkey under drought stress,
(II) to establish the relationships between morphological variations among populations and the physiological and biochemical responses of seedlings to drought stress, and
(III) to provide recommendations for the selection of populations with better adaptation to drought for future afforestation activities.
It was determined that total carotenoid, proline, and total carbohydrate contents increased, while predawn xylem water potential, chlorophyll a, chlorophyll b, and total chlorophyll amounts decreased in seedlings exposed to drought stress as the stress intensity increased. Furthermore, strong relationships were revealed between root percentage and Dickson quality index, which are important for seedling quality classification, and the physiological and biochemical responses of populations to drought stress. The populations of Alucra, Koyulhisar, and Mesudiye with water potential above -1.0 MPa during the drought stress experiment and experiencing more severe summer drought in their natural habitats have apparently exhibited higher drought tolerance compared to other populations.
Zusammenfassung
Aufgrund des Klimawandels wird erwartet, dass Waldbaumarten von Trockenheit beeinflusst werden. Da Trockenheit und Dürreperiode weltweit zu einem Problem werden, besteht die Notwendigkeit, die Reaktionen von Waldbaumarten auf Wassermangel zu untersuchen. In dieser Studie waren die Ziele (I), die physiologischen und biochemischen Reaktionen von Sämlingen zu bestimmen, die aus Samen unterschiedlicher Populationen von Quercus petraea subsp. iberica stammen, die natürlicherweise in der Türkei verbreitet sind und unter Trockenstress aufgewachsen sind, (II) Beziehungen zwischen morphologischen Variationen zwischen den Populationen und den physiologischen und biochemischen Reaktionen der Sämlinge auf Trockenstress herzustellen und (III) Empfehlungen für die Auswahl von Populationen mit besserer Anpassung an Trockenheit für zukünftige Aufforstungsaktivitäten bereitzustellen. Es wurde festgestellt, dass der Gesamt-Carotinoid-, Prolin- und Gesamtkohlenhydratgehalt zunahm, während das Xylemwasserpotential vor dem Morgengrauen, die Chlorophyll-A- und Chlorophyll-B-Werte sowie die Gesamt-Chlorophyllmengen bei Sämlingen, die Trockenstress ausgesetzt waren, mit zunehmender Stressintensität abnahmen. Darüber hinaus wurden starke Beziehungen zwischen Wurzelprozentsatz und Dickson-Qualitätsindex, die für die Klassifizierung der Sämlingsqualität wichtig sind, und den physiologischen und biochemischen Reaktionen der Populationen auf Trockenstress aufgezeigt. Die Populationen von Alucra, Koyulhisar und Mesudiye, die während des Trockenstressexperiments ein Wasserpotenzial über -1,0 MPa aufwiesen und in ihren natürlichen Lebensräumen stärkere Sommertrockenheit ausgesetzt waren, zeigten im Vergleich zu anderen Populationen eine höhere Toleranz gegenüber Trockenheit und Dürreperioden.
1 Introduction
The effects of global warming and climate change manifest as drought, decreased water resources, increased flooding, reduction of agricultural and forest areas, heatwaves, etc. (Cook et al. 2015; Turan 2018; Çobanoğlu et al. 2023). Climate change is linked to escalating aridity pressure in every forest ecosystem across the globe, with a potentially major impact on their role as carbon sinks (Anderegg et al. 2015).
It is stated that there is a correlation between a species' tolerance to drought in their native habitats and the intensity of summer droughts in those regions (Lenoir et al. 2008; Franklin et al. 2016). The identification of drought-resistant origins and their utilization in suitable growing environments are of great importance for future seed transfer and breeding programs (Ericsson et al. 1993; Dirik 2000). In nature, water stress is prevalent for either long-term or short-term periods depending on the local climate. Therefore, most plants possess some degree of adaptation or response to enhance growth and survival rates during water stress and subsequent recovery (Arve et al. 2011). Even in regions where water scarcity has not significantly affected the growth of tree species so far, problems associated with drought stress can be encountered in the near future. Therefore, it becomes important to compare the tolerances of populations within the same climatic regions in relation to drought stress (Atar 2021). This approach will also play a crucial role in establishing the foundation for active gene conservation in an uncertain future (Chaves and Oliveira 2004). Therefore, it is important to identify relatively drought-tolerant origins, as the assurance of local origins may not be certain, for determining future afforestation strategies (Çalıkoğlu 2002).
The growth and adaptation of forest trees are regulated by genetic structure and environmental factors, as in other organisms (Kozlowski and Pallardy 1997). It has been reported by various researchers that water, either alone or in conjunction with other environmental factors, is an important factor determining vegetation distribution on Earth (Kozlowski and Pallardy 1997; O'Brien 1998). Due to the significance of water in plant development, numerous studies have been conducted on water stress resulting from either water scarcity or water excess (Bayar and Deligöz 2021; Illescas et al. 2022; Koç et al. 2022; Koç and Nzokou 2023). Water stress can change the metabolic and growth patterns in the plant, reduce respiration, photosynthesis and ion absorption, and in severe situations, culminate in plant mortality (Jaleel et al. 2009).
Plants exposed to stress factors respond with physiological and biochemical reactions to prevent or minimize damage (Scholz et al. 2012). Drought stress, which is an important stress factor, triggers many biochemical, physiological, and molecular responses in plants (Cotrozzi et al. 2016; Xiong et al. 2022). As a result, plants also develop adaptation mechanisms in response to stress conditions to adapt to changing environmental conditions (Ranjan et al. 2022). In the case of drought stress, plants close their stomata and reduce carbon dioxide uptake to prevent water loss (Pirasteh‐Anosheh et al. 2016). Additionally, plants exposed to prolonged drought stress regulate their carbon assimilation rates by making certain morphological (such as leaf orientation) or physiological (such as osmotic potential) adjustments (Wright et al. 2015). Plants exposed to drought stress regulate their osmotic potentials by accumulating certain organic solutes within their cells to maintain cell turgor. These organic solutes, known also as osmolites, accumulate as soluble sugars, such as glucose and sucrose, which are soluble carbohydrates (Huang et al. 2000). Additionally, another osmolite that can be accumulated under drought stress is proline (Anjum et al. 2011). The accumulated proline under stressful conditions provides energy for plant growth and survival, aiding the plant in tolerating stress (Sankar et al. 2007). Some studies have reported an increase in soluble sugar and proline content as a result of drought stress (Deligöz and Bayar 2018; Zolfaghari and Akbarinia 2018). On the other hand, it has been stated by many researchers that drought stress has a negative impact on photosynthesis as a consequence of both stomata regulation and structural changes (Ashraf and Harris 2013; Osakabe et al. 2014). Water stress leads to structural changes in chloroplasts that affect photosynthesis (Dubey 1997). Under drought stress, the content of photosynthetic pigments (chlorophyll a, chlorophyll b, carotenoids) can decrease (Pukacki and Kaminska-Rozek 2005; Terzi et al. 2010). Following relief from drought stress and exposure to adequate water conditions, the photosynthetic systems of plants are restored (Nar et al. 2009). These responses in plants against water deficiency vary depending on genotype, species, severity and duration of water loss, plant age, and developmental stage (Clua et al. 2009).
Oaks (Quercus sp.) are known for their drought tolerance (Johnson et al. 2019). However, the level of drought tolerance may vary among different oak species (Dickson and Tomlinson 1996; Popović et al. 2010; Deligöz and Bayar 2017; Bayar 2022). Turkey has a important diversity in terms of oak species. Sessile oak (Quercus petraea), which is among the oak species with a wide distribution range, is an important species in Turkey due to its ability to naturally grow in different ecological conditions, being a tree of temperate climates, and its tolerance to low rainfall and drought (Anşin and Özkan 2006). It is also valued for its adaptability to various growing conditions and the versatility of its wood, which finds applications in many areas. The general distribution of Quercus petraea includes Europe, the Balkans, Thrace, and Anatolia. Quercus petraea subsp. iberica is naturally distributed in the Marmara region and the entire Black Sea region (Öztürk 2013). Numerous studies have been conducted on various aspects of seedling development (Chaar et al. 1997; Farque et al. 2001), fertilization (Vernay et al. 2018; Durand et al. 2020), seed germination (Kollmann and Schill 1996; Tilki 2010), forest management and regeneration (Mölder et al. 2019; Kohler et al. 2020) related to the species Quercus petraea. The effects of drought stress on sessile oak have also been revealed through various studies (Bruschi 2010; Turcsán et al. 2016; Móricz et al. 2021; Matoušková et al. 2022; Nyamjav 2022). However, there is currently no study available in Turkey that specifically investigates the drought tolerance of Quercus petraea subsp. iberica and the variations in response to drought stress among different populations. In this study, the research questions we want to address are the following: (I) determining the physiological and biochemical responses of seedlings grown from seeds obtained from different populations of Quercus petraea subsp. iberica naturally distributed in Turkey under drought stress, (II) establishing the relationships between morphological variations among populations and the physiological and biochemical responses of seedlings to drought stress, and (III) providing recommendations for the selection of Quercus petraea subsp. iberica populations with better adaptation to drought for future afforestation activities.
2 Materials and Methods
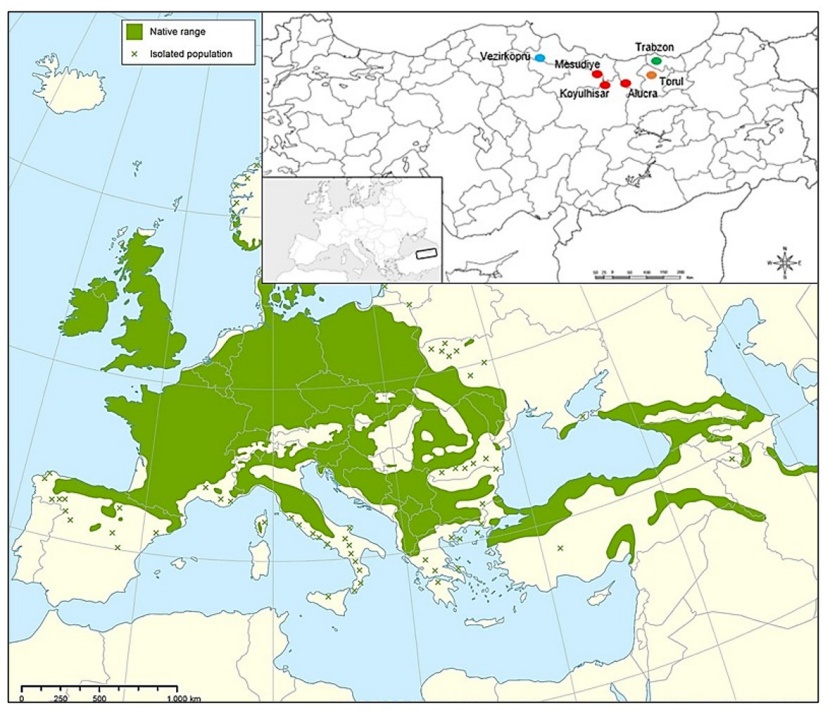
Figure 1: The geographical locations of the populations and the distribution of the groups obtained through hierarchical clustering analysis (Caudullo et al. 2017).
Abbildung 1: Die geografischen Standorte der Populationen und die Verteilung der Gruppen, ermittelt durch hierarchische Clusteranalyse (Caudullo et al. 2017).
2.1 Plant Material and Growth Conditions
The seeds were collected in September-October 2019 from six different populations of Quercus petraea subsp. iberica in the natural distribution areas in the Black Sea Region, including Trabzon-Merkez, Gümüşhane-Torul, Giresun-Alucra, Sivas-Koyulhisar, Ordu-Mesudiye, and Samsun-Vezirköprü (Table 1, Fig. 1). Healthy seeds were mostly identified by using the water flotation method, and they were then stored at cold temperature of 4°C until the sowing date. The seeds were sown in December 2019 in polyethylene bags measuring 12×20 cm, which were filled with a mixture of forest soil, peat, and river sand (2:2:1). The sowing took place in the open field nursery conditions of the Research and Application Greenhouse of the Faculty of Forestry at Karadeniz Technical University (40°59'N, 39°46'E, altitude 60 m above sea level).
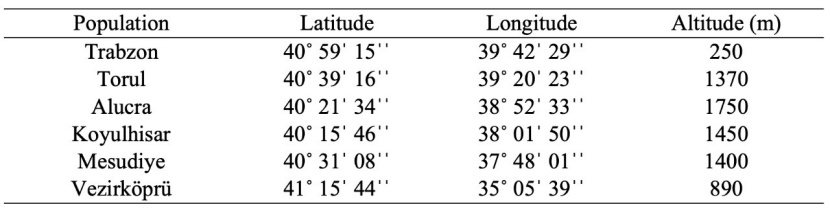
Table 1: Coordinate and altitude information for each population.
Tabelle 1: Koordinaten- und Höheninformationen für jede Population.
2.2 Experimental Design
The seedlings grown in the open-field nursery conditions were irrigated close to field capacity every two or three days from May to August 2020. In July, the seedlings were moved to the Research and Application Greenhouse to protect them from possible rainfall. Furthermore, during the implementation of the water withholding method to induce drought stress in the seedlings, the temperature in the automated greenhouse system was kept as stable as possible, and the impact of temperature fluctuations in external weather conditions was minimized. In August, the seedlings were divided into two groups for the drought stress experiment. In the control group, irrigation continued regularly (every three days until field capacity water content was reached), while irrigation was ceased in the stress group to create drought stress. The stress group was subjected to water deprivation for 5, 15, 25, 35, 45, and 55 days to create drought stress, and measurements were taken during these days. The study was conducted on a total of 1260 seedlings using a randomized complete block design with 6 populations × 2 treatments [control and drought stress] × 6 measurements × 5 seedlings × 3 replications. During the drought stress experiment, a portable meteorological station (Davis Vantage Pro-2) was installed inside the greenhouse to determine daily air temperature and humidity changes. Throughout the experiment period, the average temperature ranged between 23.4°C and 24.5°C, and the average relative humidity varied between 78.4% and 82.7%.
2.3 Predawn xylem water potential and soil moisture content
Predawn xylem water potential (PWP) was measured using a pressure chamber device (PMS Instruments, Corvallis, OR, USA) following conventional methods. Predawn measurements were conducted on nine shoot samples (three seedlings × three replications) excised from the root collar level of each population, from both the control and drought stress groups, between 03.30 and 06.00 AM. Soil moisture was measured using a soil moisture meter device (TFA Dostmann). The soil moisture meter provides values ranging from 1 to 10, with values between 1–3 indicating dry, 4–7 indicating moist, and 8–10 indicating wet conditions on the device's scale.
2.4 Photosynthetic pigments
Photosynthetic pigments (chlorophyll a, chlorophyll b, total chlorophyll and carotenoid) were determined by Arnon's method (1949). Fresh leaves from the seedlings (nine seedlings in each treatment and population), obtained by cutting them from the root collar, were cut into small pieces with scissors to achieve a homogeneous mixture. Subsequently, the fresh leaf samples weighing 0.1 g were homogenized by grinding with 10 ml of 80% acetone solution, and mixed using a vortex for approximately 5–10 seconds. The absorbance of the supernatant was measured at 450, 645 and 663 nm wavelengths using a spectrophotometer (T80+UV/VIS spectrophotometer).
2.5 Total soluble carbohydrate and proline content
Total soluble carbohydrate and proline content were measured using all of the leaves from nine seedlings in each treatment and population. Leaf samples were dried at 65˚C for 48 hours. The total soluble carbohydrate content was determined using the phenol-sulfuric acid method according to Dubois et al. (1956). An amount of 0.1 g of dried leaf samples was homogenized with 80% ethanol for 24 hours. The supernatant was transferred to another test tube and treated with a 5% phenol solution and sulfuric acid. The total carbohydrate content was calculated by creating a standard curve using standard glucose and measuring the absorbance at 490 nm, expressed as mg g-1 dry weight. The amount of proline was determined using the acid-ninhydrin method (Bates et al. 1973). An amount of 0.1 g of dried samples was homogenized with 10 ml of 3% sulfosalicylic acid. Then, 2 ml of the supernatant was taken and 2 ml of acid-ninhydrin and 2 ml of glacial acetic acid were added to it. The prepared samples were kept in a water bath at 100˚C for 1 hour and the reaction was stopped in an ice bath. Toluene was added to the cooled samples and mixed in a vortex mixer for 15 seconds, making them ready for measurement. The absorbance value of each sample was measured at 520 nm, and the proline content was calculated using a calibration curve, expressed as µmol g-1 dry weight.
2.6 Morphological parameters
Morphological measurements were conducted in October 2020 using 30 randomly selected seedlings from each population. On one-year-old seedlings, the following morphological measurements were taken: seedling length (SL), root collar diameter (RCD), shoot fresh weight (SFW), shoot dry weight (SDW), root fresh weight (RFW), root dry weight (RDW), root percentage (RP), and Dickson quality index (DQI). The seedling height was measured using a meter with a measurement accuracy of ±1 mm, and the root collar diameter was measured using a digital caliper with a measurement accuracy of ±0.01 mm. Fresh shoot and root weights were measured using a precision scale with an accuracy of ±0.001 g, and then they were dried at 105°C for 24 hours. Shoot and root dry weights were also measured using a precision scale with an accuracy of ±0.001 g. The root percentage expresses the ratio of root dry weight to the total dry weight of the seedling [RP = RDW / (SDW + RDW)]. The Dickson Quality Index was calculated using the equation DQI = (SDW + RDW) / [(SL / RCD) + (SDW / RDW)].
2.7 Climate data and bioclimatic classification for populations
Long-term climate data (1976–2000) and bioclimatic characteristics according to the Emberger bioclimatic classification for populations selected from different growing conditions are provided in Table 2. The climate data was obtained from the WorldClim database (Fick and Hijmans 2017).
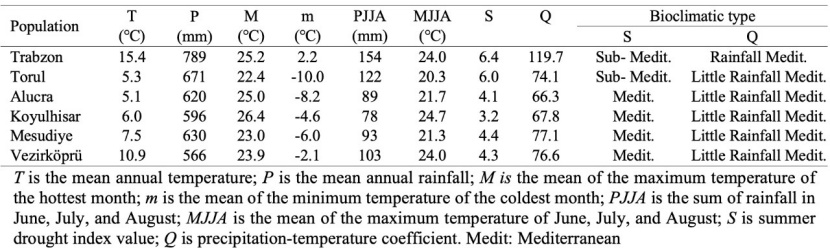
Table 2: Long-term climate data and bioclimatic types of Q. petraea subsp. iberica populations.
Tabelle 2: Langzeitklimadaten und bioklimatische Arten von Q. petraea subsp. iberica Populationen.
According to the Emberger bioclimatic classification (Akman 2011), the precipitation-temperature coefficient (Q) was applied to determine the Mediterranean bioclimatic zones, and the following formula was used. One of the most important factors in determining the Mediterranean climate is the identification of summer drought index (S). Emberger developed the following formula (Eq. 1) to determine the drought period.
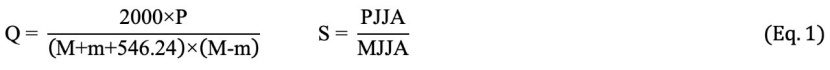
The smaller the precipitation-temperature coefficient (Q), the drier the climate, and the larger it is, the more humid the climate is. Classifications based on Q values are as follows: Q < 20: Very arid Mediterranean, 20 ≤ Q ≤ 32: Arid Mediterranean, 32 < Q ≤ 63: Semi-arid Mediterranean, 63 < Q < 98: Little-Rainfall Mediterranean, Q = 98: Rainfall Mediterranean.
As the summer drought index (S) decreases, the severity of summer drought increases, and as the summer drought index increases, the severity of summer drought decreases. Classifications based on S values are as follows: S < 5: Mediterranean S = 5–7: Sub-Mediterranean S > 7: Not Mediterranean.
2.8 Statistical analysis
The data were analyzed using the “Windows SPSS Software 26.0” and “R v.4.1.3” statistical package programs. Variance analysis was applied to determine the statistical significance (p<0.05) of the differences in morphological, physiological, and biochemical characteristics among populations. Independent samples T-test was used to determine the statistical significance (p<0.05) of differences between drought stress treatments for each measurement day. Correlation analysis was performed to reveal the linear relationships between the measured morphological, physiological and biochemical characteristics. The results of the morphological, physiological, and biochemical characteristics were evaluated together with the long-term climate data of the populations, and the groupings formed among the populations were determined using hierarchical clustering analysis. The statistical significance of the grouping obtained by clustering analysis was tested using discriminant analysis.
3 Results
Statistically significant differences (p<0.01) were determined among the measurement time, drought stress application and populations for all measured physiological and biochemical parameters. Furthermore, significant differences at a 99% confidence level were determined among each measured parameter depending on the interactions of time × stress, time × population, stress × population, and time × stress × population (Table 3).
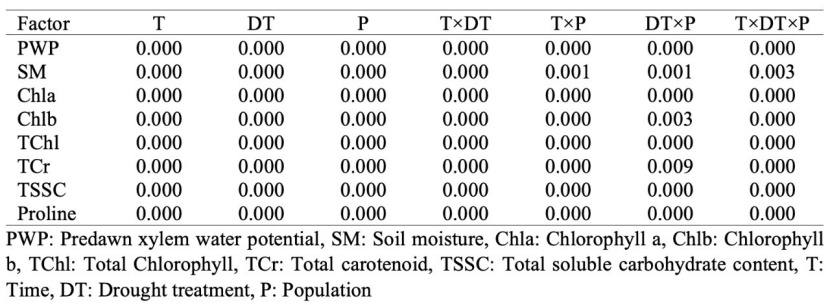
Table 3: Results of the variance analysis for the measured parameters.
Tabelle 3: Ergebnisse der Varianzanalyse für die gemessenen Parameter.
3.1 Drought stress impacts on predawn xylem water potential and soil moisture content
Predawn xylem water potential values were found to vary depending on drought stress application and populations. The average PWP values of the populations included in the control group did not show much difference depending on the measurement time. In the control group, the average PWP value was measured highest at -0.15 MPa (Vezirköprü population, measurement on the 15th day) and lowest at -0.42 MPa (Koyulhisar population, measurement on the 55th day). In the drought group, the highest average PWP value was determined as -0.22 MPa (Torul population, measurement on the 5th day), and the lowest average PWP value was -2.13 MPa (Trabzon population, measurement on the 55th day). In Trabzon, Torul, and Vezirköprü populations, it was determined that starting from the 45th day of the drought stress experiment, the average PWP dropped below -1.00 MPa, and yellowing and drying began on the leaves of some seedlings. 55th day, PWP values dropped below -2.00 MPa, and most of the seedlings dried out. The average PWP values of the Alucra, Koyulhisar, and Mesudiye populations did not drop below -1.10 MPa even on the 55th day of drought stress, indicating that their drought tolerance is higher compared to other populations.
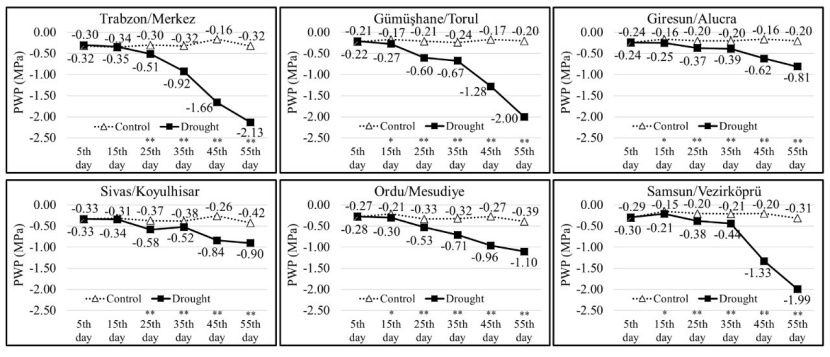
Figure 2: PWP values and t-test results (*p<0.05, **p<0.01) for measurement days and populations.
Abbildung 2: PWP-Werte und t-Test-Ergebnisse (*p<0.05, **p<0.01) für Messtage und Populationen.
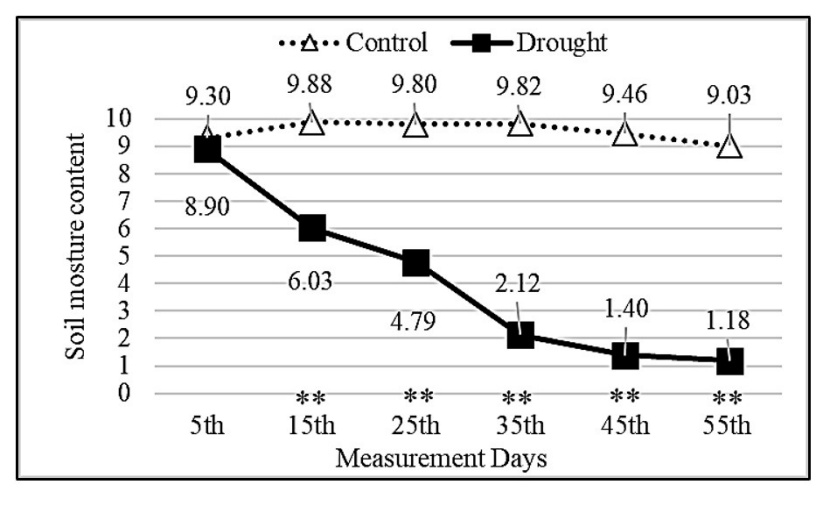
Figure 3: Change of soil moisture depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 3: Veränderung der Bodenfeuchtigkeit in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
Until the 15th measurement day, the PWP values were close to each other in the control and drought groups. However, the PWP values decreased as the water deficiency increased, starting on the 25th measurement day in the drought group. As seen in Fig. 3, soil moisture levels in the control group were generally close to each other as a result of regular irrigation, while soil moisture levels in the drought group decreased constantly due to increased drought stress. As a result of the independent samples t-test, statistically significant differences at a 99% confidence level were found on each measurement day except for the 5th measurement day.
3.2 Drought stress impacts on photosynthetic pigments
The amounts of chlorophyll a, chlorophyll b and total chlorophyll generally showed fluctuating course depending on the measurement times in the control group. However, the chlorophyll levels in the drought group were lower compared to the control group starting on the 25th measurement day. According to the independent samples t-test results, statistically significant differences (p<0.05) were found between the results obtained on the 35th and 55th measurement days of the control and drought groups. The total carotenoid amounts of the drought group had higher values compared to the control group starting on the 15th measurement day. However, no statistically significant differences (p>0.05) were found between the results obtained from the control and drought groups on each measurement day (Fig. 4).
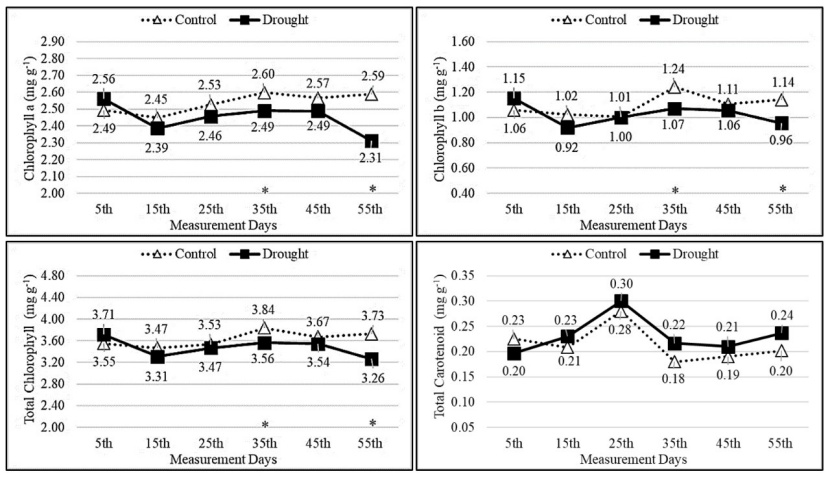
Figure 4: Change of chlorophyll a, chlorophyll b, total chlorophyll and total carotenoid amounts depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 4: Veränderung der Mengen an Chlorophyll a, Chlorophyll b, Gesamtchlorophyll und Gesamt Karotinoide in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
In the control group, the minimum TChl was determined to be 2.60 mg g-1 (on the 15th measurement day in the Torul population), and the maximum TChl was 4.81 mg g-1 (on the 35th measurement day in the Vezirköprü population). In the drought group, the minimum TChl was 1.92 mg g-1 (on the 55th measurement day in the Trabzon population), and the maximum TChl was 4.54 mg g-1 (on the 45th measurement day in the Koyulhisar population). As the measurement days increased in the Trabzon, Torul, Mesudiye, and Vezirköprü populations, the total chlorophyll amounts of the seedlings belonging to the drought group generally remained lower compared to the control group. However, in the Alucra and Koyulhisar populations, it was observed that the total chlorophyll levels varied depending on the measurement days for both the control and drought groups (Fig. 5).
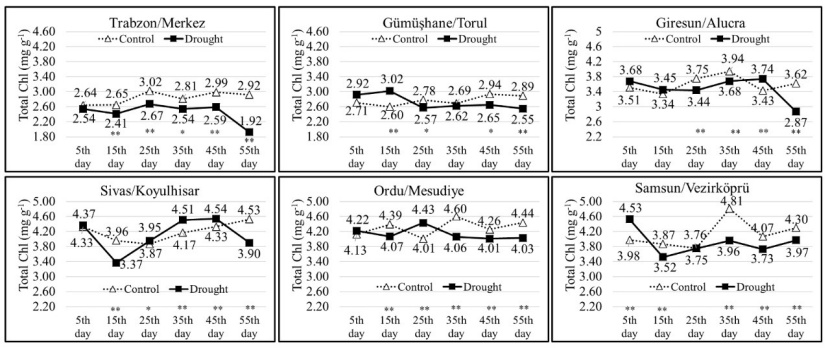
Figure 5: Change of total chlorophyll amounts in each population depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 5: Veränderung der gesamten Chlorophyllmengen in jeder Population in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
3.3 Drought stress impacts on total soluble carbohydrate and proline content
In the control group, the total soluble carbohydrate and proline contents showed a close and gradually increasing trend depending on the measurement days. The drought group generally had close values to the control group until the 25th measurement day, but the total soluble carbohydrate and proline contents of the seedlings in the drought group increased from the 35th day with increasing drought stress (Fig. 6).
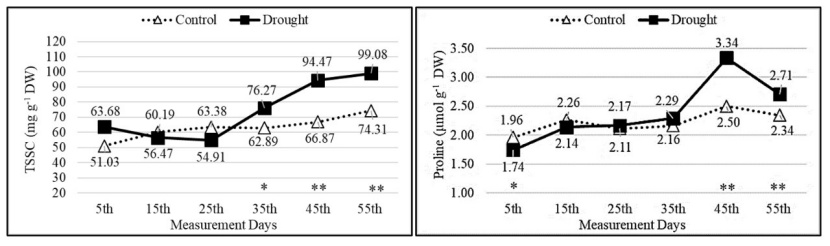
Figure 6: Change of total soluble carbohydrate and proline contents depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 6: Veränderung des Gesamtgehalts an löslichen Kohlenhydraten und Prolingehalt in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
The TSCC of the populations in the drought group showed significant changes throughout the duration of drought stress. In the drought group, the TSCC was the lowest at 43.15 mg g-1 (on the 15th measurement day in the Torul population) and the highest at 122.31 mg g-1 (on the 55th measurement day in the Trabzon population). Until the 35th measurement day, the TSCC of the seedlings in the Trabzon, Torul, Alucra and Vezirköprü populations were close to each other in the control and drought groups. However, from the 35th day onward, the TSCC of the seedlings in the drought group increased due to the effect of drought stress. In both Koyulhisar and Mesudiye populations, the TSCC of the drought and control groups exhibited variation throughout the drought stress application, and on the 55th measurement day, the TSCC of the seedlings in the drought group was higher compared to the seedlings in the control group (Fig. 7). Except for the Mesudiye population, in other populations, as the duration of drought stress application increased, the seedlings in the drought group had higher proline contents compared to the seedlings in the control group. The seedlings exposed to drought stress accumulated proline as the stress increased (Fig. 8).
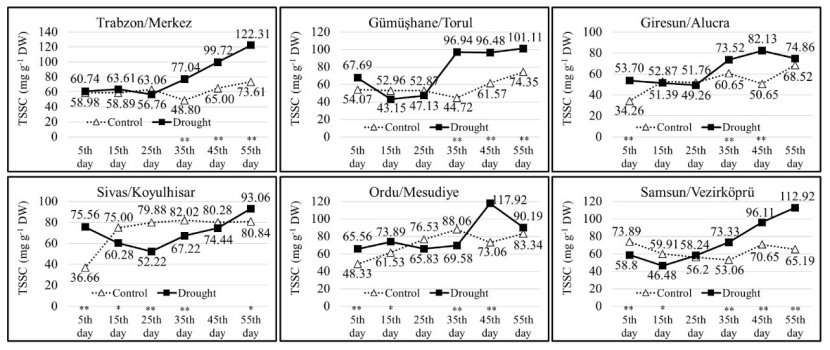
Figure 7: Change of total soluble carbohydrate content in each population depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 7: Veränderung des Gesamtgehalts an löslichen Kohlenhydraten in jeder Population in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
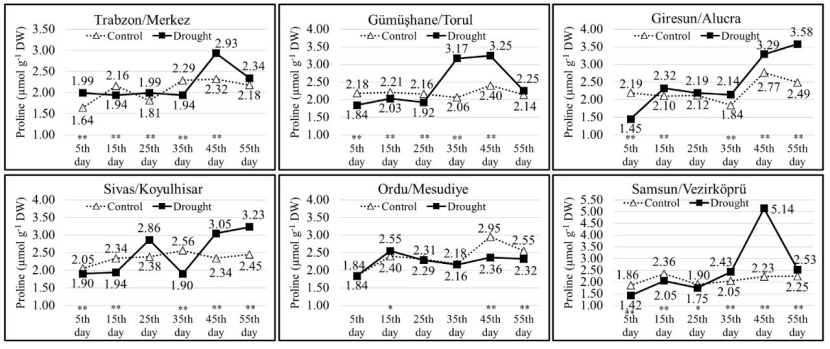
Figure 8: Change of proline content in each population depending on measurement days and treatments. Asterisks show significant differences by independent sample t-test (*p< 0.05, **p< 0.01) between treatments.
Abbildung 8: Veränderung des Prolingehalts in jeder Population in Abhängigkeit von Messtagen und Behandlungen. Sternchen zeigen signifikante Unterschiede zwischen den Behandlungen anhand des unabhängigen Stichproben-T-Tests (*p<0.05, **p<0.01).
3.4 Drought stress impacts on morphological parameters
In order to reveal the relationship between the responses of seedlings from different populations to drought stress and their morphological characteristics, some morphological characteristics of the seedlings were determined. Based on the results of the analysis of variance, there was a statistically significant difference among populations at the 99% confidence level in terms of SL, SFW, SDW, RP, and DQI values. In terms of RCD and RDW values, there was a statistically significant difference among populations at the 95% confidence level. However, there was no statistically significant difference among populations in terms of RFW value (Table 4).
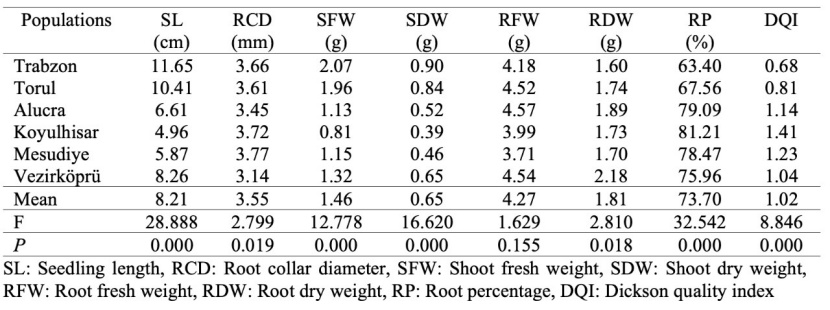
Table 4: Mean values and analysis of variance results for morphological characteristics of populations.
Tabelle 4: Mittelwerte und Varianzanalyseergebnisse für morphologische Merkmale von Populationen.
The significance of the groups formed among populations through hierarchical cluster analysis was tested using discriminant analysis, and the separation into two groups was found to be statistically significant. According to Emberger's precipitation-temperature coefficient (Q), the populations of Alucra, Mesudiye, Koyulhisar, Vezirköprü, and Torul, located in the low-rainfall Mediterranean Bioclimatic type, form the first group, while the population of Trabzon, located in the rainfall Mediterranean Bioclimatic type, forms the second group (Fig. 9). Although the discriminant analysis determined that the separation into two groups was statistically significant, the other potential groups that could be formed are shown in Fig. 1 and 9. Based on S and Q values (Table 2), the populations of Alucra, Mesudiye, and Koyulhisar, which have low values, were in the first group. The population of Vezirköprü formed another group. The populations of Torul and Trabzon, which have higher S and Q values compared to the other populations, were also in separate groups on their own. Indeed, it can be observed from the clustering that the climatic characteristics of populations and consequently their adaptation to their habitats have a significant influence on the morphological characteristics as well as the physiological and biochemical responses to drought stress.
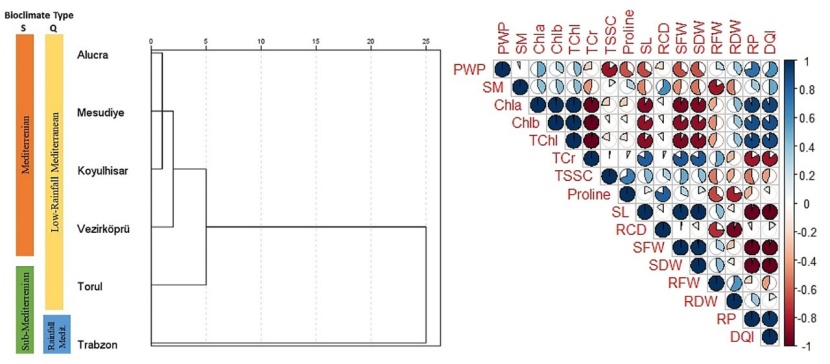
Figure 9: Dendrogram of population groups related to hierarchical cluster analysis, and correlation analysis result of measured morphological, physiological and biochemical parameters.
Abbildung 9: Dendrogramm der Bevölkerungsgruppen im Zusammenhang mit der hierarchischen Clusteranalyse und Korrelationsanalyseergebnis der gemessenen morphologischen, physiologischen und biochemischen Parameter.
According to the correlation analysis, it was determined that the PWP value has a positive correlation with Chla, Chlb, and TChl, while it has a negative correlation with TSCC and proline. Furthermore, the PWP value is strongly positively correlated with the RP and DQI of the seedlings, while it has a negative relationship with SL, SFW and SDW. It has been revealed that many measured parameters exhibit strong correlations, both negative and positive, with each other (Fig. 9). Indeed, the correlation analysis has showed that there are strong relationships between the physiological and biochemical responses of the seedlings to drought stress and their morphological characteristics.
Statistically significant relationships were found, both negative and positive, between climate data and the measured characteristics. The S and Q values, used in Emberger's climate classification, were positively correlated with TCr, TSCC, SL, SFW and SDW and negatively correlated with PWP, Chla, Chlb, TChl, RP and DQI (Table 5). Populations with low summer drought indexes (S) and rainfall-temperature coefficients (Q) had higher PWP values because they lost less water during drought stress due to their adaptation to the growing environments.
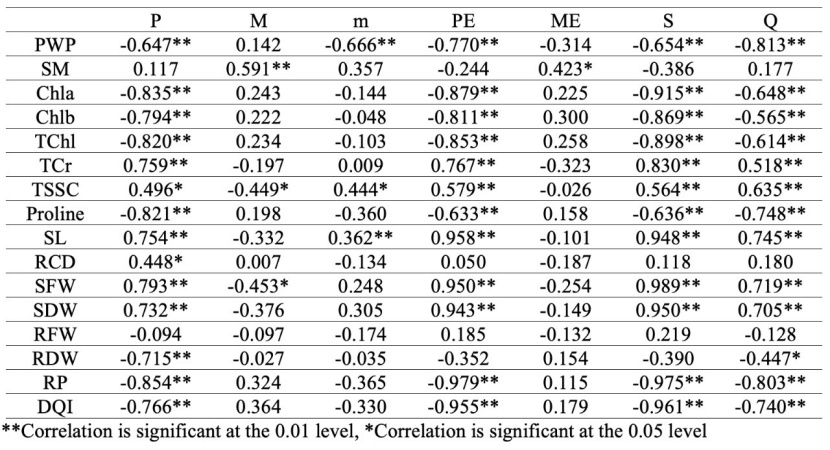
Table 5: Correlation analysis result of climate data and measured parameters.
Tabelle 5: Ergebnis der Korrelationsanalyse von Klimadaten und gemessenen Parametern.
4 Discussion
4.1 Drought stress impacts on physiological and biochemical characteristics
Plants reach their highest water potential values during the predawn (Cleary and Zaerr 1984). In many studies, it is indicated that the optimal PWP for plant growth is between -0.5 and -1.2 MPa, and when the PWP drops below -1.2 MPa, the plant requires irrigation. (McDonald 1984; Lopushinsky 1990). In the present study, at the beginning of the drought stress experiment, the highest average PWP was obtained as -0.22 MPa in the Torul population, while on the 55th day of the drought stress experiment, the lowest average PWP was determined as -2.13 MPa in the Trabzon population. Deligöz and Bayar (2017) found in their study on Q. cerris that the xylem water potential, which was -0.84 MPa at the beginning of the drought stress experiment, decreased to -1.73 MPa on the 30th day of the experiment in stressed seedlings. Also, in the control seedlings, the plant water potentials ranged from -0.56 MPa to -1.01 MPa. Thomas and Gausling (2000) stated that under moderate drought stress, Q. petraea and Q. robur seedlings had significantly lower predawn leaf water potentials compared to control seedlings. In a study conducted on Q. variabilis, it was reported that leaf water potential values decreased as drought stress increased (Wu et al. 2013). As a result of drought stress applied to Q. cerris and Q. robur seedlings, the lowest predawn xylem water potential was determined as -2.28 MPa for Q. cerris and -3.37 MPa for Q. robur (Deligöz and Bayar 2018). Consistent with previous research results, our study also revealed a significant decrease in xylem water potentials during the duration of drought stress.
Plant water potential values can vary depending on environmental conditions and plant species. Indeed, significant differences were observed among populations in terms of PWP values in the present study. In a study performed on seedlings belonging to 7 different origins representing different bioclimatic zones of Pinus brutia, it was determined that origins experiencing strong summer drought in their natural distribution areas had the lowest osmotic potential values and were the most resistant to drought effects (Dirik 2000). A seedling could survive at a predawn water potential of -2.0 MPa, but typical stomatal activities may be inhibited during the day, photosynthesis may progressively decline, and seedling development may cease (Lopushinsky 1990). In the present study, it has been determined that in particular populations of Trabzon, Torul, and Vezirköprü, the average PWP dropped below -1.0 MPa from the 45th day of the drought stress experiment. Some seedlings started showing yellowing and wilting symptoms. The PWP had dropped below -2.0 MPa by the 55th day, and many seedlings experienced severe wilting. On the other hand, in the populations of Alucra, Koyulhisar, and Mesudiye, it was observed that the average PWP remained above -1.0 MPa even on the 55th day of the drought stress, indicating their high drought tolerance.
Changes in light intensity and the amount of water in the soil significantly affect the synthesis of pigments (Matysiak 2001). This is because chlorophyll pigments are highly sensitive to various environmental factors (Lepeduš et al. 2003). It is known that carotenoids protect the photosynthetic membranes from damage caused by light and play a role in capturing photosynthetic light (Havaux 1998). In this study, it was found that from the 25th measurement day onwards, the drought group exhibited lower amounts of chlorophyll a, chlorophyll b, and total chlorophyll compared to the control group, while the total carotenoid amounts were higher. Additionally, positive correlations were observed between the PWP and chlorophyll amounts, whereas a negative correlation was found with total carotenoid. Similarly, Baquedano and Castillo (2007) reported a significant reduction in chlorophyll amounts in oak species due to the impact of drought stress. Mafakheri et al. (2010) pointed out that drought stress applied during plant growth significantly reduced the amounts of chlorophyll a, chlorophyll b, and total chlorophyll.
One of the responses of plants to drought stress is the alteration of carbohydrates in stressed plants (Vassey and Sharkey 1989). In response to drought conditions, carbohydrate accumulation increases in different parts of the plant (Krasenky and Jonak 2012). In the present study, the Trabzon, Torul, and Vezirköprü populations, which were significantly affected by water insufficiency and whose PWP values approached and fell below -2.0 MPa by the 55th measurement day, had higher carbohydrate accumulation compared to other populations. Similar to the results of the study, Deligöz and Bayar (2017) emphasized that starting from the 7th day of the drought experiment in Q. cerris, the total soluble carbohydrate content in drought-stressed seedlings was found to be significantly higher compared to the control seedlings. In studies conducted on Q. pubescens (Holland et al. 2016) and Q. variabilis (Wu et al. 2013) seedlings, it was found that the total soluble carbohydrate content increased with the increase in drought stress level. Many studies have also reported carbohydrate accumulation in plants under drought stress conditions (Morales et al. 2013; Maguire and Kobe 2015; Deligöz and Bayar 2018).
One of the most important responses of plants to drought stress is the accumulation of protective solutes such as proline (Yavaş et al. 2016). Many plants accumulate proline as a response to osmotic stress (Bhaskara et al. 2015). The concentration of proline increases with a decrease in water potential or an increase in plant water stress (Lansac et al. 1994). In the present study, it was determined that the proline amount of the seedlings in the drought group was higher than the seedlings in the control group due to the increase in drought stress in other populations except the Mesudiye population. It is believed that the lower accumulation of proline in the drought group of the Mesudiye population, compared to the control group, is due to the prioritization of carbohydrate accumulation rather than proline accumulation by the seedlings under drought stress to maintain osmotic balance. Indeed, when examining the results of carbohydrate analysis, it can be seen that the Mesudiye population had the highest carbohydrate accumulation among the populations with a value of 117.92 mg g-1 on the 45th measurement day. Cotrozzi et al. (2016) reported that drought stress caused an increase in proline amounts in Q. ilex, Q. pubescens, and Q. cerris species. Deligöz and Bayar (2018) stated that drought stress induced an increase in proline and carbohydrate contents in Q. cerris and Q. robur, and Q. robur had more proline accumulation than Q. cerris. Wu et al. (2013) indicated that drought stress increased proline amounts in Q. variabilis seedlings. Numerous studies have also observed an increase in proline amounts with the increase in drought stress (Sircelj et al. 2005; Shvaleva et al. 2006; Yang et al. 2007).
4.2 Drought stress impacts on morphological characteristics
It is seen that many of the studies conducted until recent years for determining seedling quality were based on morphological parameters (Apholo and Rikala 2003; Atar 2021; Güney et al. 2023). In a study conducted by Semerci (2002), a positive correlation was found between seedling length and survival success. However, some researchers have reported a negative correlation between these two parameters (Larsen et al. 1986; Tuttle et al. 1987). Furthermore, it is stated that the root collar diameter is a more important criterion in seedling quality classification compared to seedling length (Yahyaoğlu and Genç 2007). The root percentage is another important parameter used to determine seedling quality (Ayan et al. 2020; Güney et al. 2020). High root development potential is associated with a high survival rate in the field (Ritchie 1984). Among the populations, the highest mean RP (81.21%) was determined in the Koyulhisar population, which is one of the populations least affected by drought stress, and the lowest mean RP (63.40%) was determined in the Trabzon population, which was most affected by drought stress. Bruschi (2010) stated that there are significant differences in morphological characteristics in Q. petraea seedlings exposed to drought stress, both in terms of population and irrigation regime. The Dickson quality index is an important indicator in determining seedling planting success and survival rate (Bayala et al. 2009). According to Dickson et al. (1960), seedlings with a quality index close to or above 1 are considered to be of higher quality. In this study, the highest mean DQI was obtained in the Koyulhisar population with a value of 1.41, while the lowest mean DQI was found in the Trabzon population with a value of 0.68. The populations with the highest DQI values, namely Koyulhisar, Alucra, and Mesudiye, were determined to be the most drought-tolerant populations based on their responses to drought stress. In addition, it was determined that there was a strong positive correlation between PWP and RP and DQI values of the seedlings by correlation analysis. Indeed, Ritchie and Shula (1984) emphasized that evaluating the physiological characteristics of seedling material is crucial, regardless of how good they are in terms of genetic and morphological features, as accurate results cannot be obtained otherwise. Furthermore, they reported that the results would be more accurate when the morphological data used for seedlings aligned with physiological data.
5 Conclusions
In order to make afforestation efforts successful in arid and semi-arid areas, knowing the phenotypic characteristics of existing forest trees in nature may not always be sufficient. Therefore, it is important to determine the geographic variations of species and assess the morphological and physiological characteristics of seeds and seedlings obtained from these areas. The populations of Alucra, Koyulhisar, and Mesudiye, which have PWP above -1.0 MPa during the drought stress experiment and experience more severe summer drought in their natural habitats (with lower S and Q values), have exhibited higher drought tolerance compared to other populations. It can be concluded that these populations can be used in afforestation efforts in water-deficient arid and semi-arid regions.
The increase in the total soluble carbohydrate content of the populations of Q. petraea subsp. iberica under stress conditions can be expressed as an adaptation developed in response to adverse conditions. The increase in both proline and total soluble carbohydrate content serves as important criteria indicating drought tolerance in Q. petraea subsp. iberica. Moreover, the increase in carotenoid amounts can be considered a good adaptation criterion for drought tolerance, as it helps prevent membrane damage associated with water deficiency and contributes to the continuity of photosynthesis by assisting chlorophyll pigments.
In the study, it has been revealed that there are strong and significant relationships between the data obtained from the root percentage and Dickson quality index, which are important parameters in the seedling quality classification, and the drought stress resistance of the populations. Thus, it can be said that morphological parameters such as RP and DQI, which are simpler to measure than physiological and biochemical parameters, can be used in determining the drought tolerance of different populations. Additionally, the bioclimatic classification created using the summer drought index (S) and precipitation-temperature coefficient (Q) can be utilized as an indicator in the selection of populations that may exhibit drought tolerance. Also, it is crucial to integrate knowledge about intraspecific ecological amplitude and adaptation processes. However, there might still be biases and difficulties in unraveling the correlation between the environment and genetics.
The fact that there are statistically significant differences in terms of drought stress among the studied populations necessitates a more detailed study by considering all distribution areas of Q. petraea subsp. iberica in Turkey. In this context, future studies to establish in situ and ex-situ protection strategies for the species and the determination of origins that are highly adaptable to changes that may occur in the horizontal and vertical distribution areas of the species, especially at the point of adaptation to climate change, will reveal extremely important results for forestry strategies.
Acknowledgements
This study was a part of the PhD thesis prepared by Graduate School of Natural and Applied Sciences at Karadeniz Technical University. Also, we would like to thank Research Assistant Ali Bayraktar for his contributions to the laboratory works.
Funding
This work was funded by Office of Scientific Research Projects of Karadeniz Technical University. Project number: FHD-2020-8863.
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
References
Akman Y (2011) İklim ve biyoiklim. Palme Yayıncılık, Ankara, Turkey.
Anderegg WR, Ballantyne AP, Smith WK, Majkut J, Rabin S, Beaulieu C, Birdsey R, Dunne JP, Houghton RA, Myneni RB, Pan Y, Sarmiento JL, Serota N, Shevliakova E, Tans P, Pacala SW (2015) Tropical nighttime warming as a dominant driver of variability in the terrestrial carbon sink. Proceedings of the National Academy of Sciences 112(51): 15591-15596.
Anjum SA, Xie X, Wang L, Saleem MF, Man C, Lei W (2011) Morphological, physiological and biochemical responses of plants to drought stress. African Journal of Agricultural Research 6: 2026-2032.
Anşin R, Özkan CÖ (2006) Tohumlu bitkiler (Spermatophytha) odunsu taksonlar. KTÜ Orman Fakültesi Genel Yayın No: 167, Fakülte Yayın No: 19, Trabzon, Turkey.
Apholo P, Rikala R (2003) Field performance of silver-birch planting-stock grown at different spacing and in containers of different volume. New Forests 25: 93-108.
Arnon DI (1949) Copper Enziymes in Izolated Chloroplasts Polyphenoioxidase in Beta vulgaris, Plant Physiolgy, 24, 1-15.
Arve LE, Torre S, Olsen JE, Tanino KK (2011) Stomatal responses to drought stress and air humidity. In abiotic stress in plants-Mechanisms and adaptations (ed. by A Shanker and B Venkateswarlu) IntechOpen, London, UK.
Ashraf M, Harris PJC (2013) Photosynthesis under stressful environments: An overview. Photosynthetica 51: 163-190.
Atar F (2021) Effects of altitude on some seedling quality characteristics of Carpinus betulus L. (common hornbeam) and Carpinus orientalis Mill. (oriental hornbeam). Artvin Çoruh Üniversitesi Orman Fakültesi Dergisi 22(2): 257-265.
Ayan S, Civek E, Yer Çelik E, Gülseven O, Akın Ş, Yılmaz E (2020) Akdeniz ve Arizona servisi fidanlarının morfolojik kalite özellikleri. Bartın Orman Fakültesi Dergisi 22(2): 580-590.
Baquedano FJ, Castillo FJ (2007) Drought tolerance in the Mediterranean species Quercus coccifera L., Quercus ilex L., Pinus halepensis Mill., and Juniperus phoenicea L. Photosynthetica 45(2): 229-238.
Bates LS, Waldren RP, Teare ID (1973) Rapid Determination of free proline for water stress studies. Plant and Soil 39: 205–207.
Bayala J, Dianda ZM, Wilson ZJ, Ouedraogo SJ, Sanon ZK (2009) Predicting field performance of five irrigated tree species using seedling quality assessment in burkina faso, West Africa. New Forests 38: 309-322.
Bayar E (2022) Influence of drought stress and N addition on the gas exchange, biochemical and growth traits in Quercus ithaburensis. Dendrobiology 88: 94-104.
Bayar E, Deligöz A (2021) Ecophysiological behavior of Mediterranean woody species under summer drought. Revista Bosque 42(3): 311-321.
Bhaskara GB, Yang TH, Verslues PE (2015) Dynamic proline metabolism: Importance and regulation in water limited environments. Frontiers in Plant Science 6: 484.
Bruschi P (2010) Geographical variation in morphology of Quercus petraea (Matt.) Liebl. as related to drought stress. Plant Biosystems 144(2): 298-307.
Caudullo G, Welk E, San-Miguel-Ayanz J (2017) Chorological maps for the main European woody species. Data in Brief 12, 662–666.
Chaar H, Colin F, Collet C (1997) Effects of environmental factors on the shoot development of Quercus petraea seedlings A methodological approach. Forest Ecology and Management 97(2): 119-131.
Chaves MM, Oliveira MM (2004) Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. J Exp Bot 55(407): 2365-2384.
Cleary BD, Zaerr JB (1984) Guidelines for measuring plant moisture stress with a pressure chamber. PMS Instrument Co., 2750 N. W. Royal Oaks Drive, Corvallis, Oregon, USA.
Clua A, Paez M, Orsini H, Beltrano J (2009) Incidence of drought stress and rewatering on Lotus tenuis effects on cell membrane stability. Lotus Newsletter 39(1):21-27.
Cotrozzi L, Remorini D, Pellegrini E, Landi M, Massai R, Nali C, Guidi L, Lorenzini G (2016) Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiologia plantarum 157(1): 69-84.
Çalıkoğlu M, Tilki F (2002) Orman ağacı tohumlarında çimlenme-su stresi ilişkisi, İÜ Orman Fakültesi Dergisi 76-88.
Çobanoğlu H, Cantürk U, Koç İ, Kulaç Ş, Sevik H (2023). Climate change effect on potential distribution of Anatolian chestnut (Castanea sativa mill.) in the upcoming century in Turkey. Forestist 73(3): 247-256.
Cook BI, Ault TR, Smerdon JE (2015) Unprecedented 21st century drought risk in the American Southwest and Central Plains. Science Advances 1(1): e1400082.
Deligöz A, Bayar E (2017) Variations in physiological and biochemical traits of drought-stressed Quercus cerris seedlings. Turkish Journal of Forestry 18(4): 269-274.
Deligöz A, Bayar E (2018) Drought stress responses of seedlings of two oak species (Quercus cerris and Quercus robur). Turkish Journal of Agriculture and Forestry 42(2): 114-123.
Dickson R, Tomlinson PT (1996) Oak growth, development and carbon metabolism in response to water stress. Ann For Sci 53: 181-196.
Dickson A, Leaf AL, Hosner F (1960) Quality appraisal of white spruce and white pine seedling stock in nurseries. Forestry Chronicle 36: 10-13.
Dirik H (2000) Farklı biyoiklim kuşaklarını temsil eden kızılçam (Pinus brutia Ten.) orijinlerinin kurak dönemdeki su potansiyellerinin basınç-hacim (p-v) eğrisi yöntemi ile analizi. İÜ Orman Fak. Dergisi Seri A 50(2): 93-130.
Dubey RS (1997) Photosynthesis in plants under stressful conditions. Handbook of photosynthesis (ed. by M Pessarakli) Dekker, New York.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Calorimetric method for determination of sugars and related substances. Analytical Chemistry 28: 350-356.
Durand M, Rose C, Dupouey JL, Legout A, Ponton S (2020) Do tree rings record changes in soil fertility? Results from a Quercus petraea fertilization trial. Science of the Total Environment 712: 136148.
Ericsson G, Namkoong G, Roberds JH (1993) Dynamic gene conservation for uncertain futures. Forest Ecology and Management 62(1-4): 15-37.
Farque L, Sinoquet H, Colin F (2001) Canopy structure and light interception in Quercus petraea seedlings in relation to light regime and plant density. Tree physiology 21(17): 1257-1267.
Fick SE, Hijmans RJ (2017) WorldClim 2: new 1‐km spatial resolution climate surfaces for global land areas. International journal of climatology 37(12): 4302-4315.
Franklin J, Serra-Diaz JM, Syphard AD, Regan HM (2016) Global change and terrestrial plant community dynamics. Proceedings of the National Academy of Sciences 113(14): 3725-3734.
Güney D, Bayraktar A, Atar F, Turna İ (2020) Effects of root undercutting, fertilization and thinning on seedling growth and quality of oriental beech (Fagus orientalis Lipsky) seedlings. Artvin Coruh University Journal of Forestry Faculty 21(2): 214-222.
Güney D, Atar F, Turna İ, Bayraktar A (2023) Morphogenetic variations of Pinus sylvestris L. seedlings depending on altitude. Turkish Journal of Forestry Research 10(1): 80-89.
Havaux M (1998) Carotenoids As membrane stabilisers in chloroplasts. Trends Plant Sci. 3: 147-151.
Holland V, Koller S, Lukas S, Brüggemann W (2016) Drought and frost-induced accumulation of soluble carbohydrates during accelerated senescence in Quercus pubescens. Trees 30(1): 215-226.
Huang XM, Huang HB, Gao FF (2000) The growth potential generated in citrus fruit under water stress and its relevant mechanisms. Scientia Horticulturae 83(3-4): 227-240.
Illescas M, Morán-Diez ME, Martínez de Alba ÁE, Hermosa R, Monte E (2022) Effect of Trichoderma asperellum on wheat plants’ biochemical and molecular responses, and yield under different water stress conditions. International Journal of Molecular Sciences 23(12): 67-82.
Jaleel CA, Manivannan P, Wahid A, Farooq M, Al-Juburi HJ, Somasundaram R, Panneerselvam R (2009) Drought stress in plants: a review on morphological characteristics and pigments composition. Int. J. Agric. Biol 11(1): 100-105.
Johnson PJ, Shifley SR, Rogers R, Dey DC, Kabrick JM (2019) The ecology and silviculture of oaks. 3rd ed. CABI Publishing, Boston, USA.
Koç İ, Nzokou P (2023) Combined effects of water stress and fertilization on the morphology and gas exchange parameters of 3-year-old Abies fraseri (Pursh) Poir. Acta Physiologiae Plantarum, 45(3): 49. doi: 10.1007/s11738-023-03529-4.
Koç İ, Nzokou P, Cregg B (2022) Biomass allocation and nutrient use efficiency in response to water stress: Insight from experimental manipulation of balsam fir, concolor fir and white pine transplants. New Forests 53(5): 915-933.
Kohler M, Pyttel P, Kuehne C, Modrow T, Bauhus J (2020) On the knowns and unknowns of natural regeneration of silviculturally managed sessile oak (Quercus petraea (Matt.) Liebl.) forests-a literature review. Annals of Forest Science 77: 1-19.
Kollmann J, Schill HP (1996) Spatial patterns of dispersal, seed predation and germination during colonization of abandoned grassland by Quercus petraea and Corylus avellana. Vegetatio 125: 193-205.
Kozlowski TT, Pallardy SG (1997) Growth control in woody plants. Academic Press, San Diaogo, USA.
Krasenky J, Jonak C (2012) Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. Journal of Experimental Botany 63(4): 1593-1608.
Lansac AR, Zaballos JP, Martin A (1994) Seasonal water potential changes and proline accumulation in Mediterranean shrubland species. Vegetatio 113(2): 141-154.
Larsen HS, South DB, Boyer JM (1986) Root growth potential, seedling morphology and bud dormancy carreiate with survival of lob-lolly pine seedlings planted in December in Alabama. Tree Physiology 1(41): 253-263.
Lenoir J, Gégout JC, Marquet, PA, de Ruffray P, Brisse H (2008) A significant upward shift in plant species optimum elevation during the 20th century. Science, 320(5884): 1768-1771.
Lepeduš H, Cesar V, Suver M (2003) The annual changes of choloroplast pigments content in current-and previous-year needles of norway spruce (Picea abies L. Karst.) exposed to cement dust pollution. Acta Botanica Croatica 62(1): 27-35.
Lopushinsky W (1990) Seedling moisture status. (ed. by R Rose, SJ Campbell and TD Landis) Target Seedling Symposium. Proceedings, Western Forest Nursey Association; August, Roseburg. OR. General Technical Report RM-200, 123-138.
Mafakheri A, Siosemardeh A, Bahramnejad B, Struik PC, Sohrabi Y (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Australian Journal of Crop Science 4(8): 580-585.
Matoušková M, Urban J, Volařík D, Hájíčková M, Matula R (2022) Coppicing modulates physiological responses of sessile oak (Quercus petraea Matt. Lieb.) to drought. Forest Ecology and Management 517: 120253.
Matysiak R (2001) Content of Carotenoids in needles of Pinus sylvestris L. growing in a polluted area. Dendrobiology 46: 39-42.
McDonald SE (1984) Irrigation in Forest-Tree Nurseries: Monitoring and Effects on Seedling Growth. Forest Nursery Manuel (ed. by ML Duryea and TD Landis) Martinus Nijhoff/DrW. Junk Publishers, Hague, Netherlands.
Morales CG, Pino MT, del Pozo A (2013) Phenological and physiological responses to drought stress and subsequent rehydration cycles in two raspberry cultivars. Scientia Horticulturae 162: 234-241.
Móricz N, Illés G, Mészáros I, Garamszegi B, Berki I, Bakacsi Z, Kampel J, Szabo O, Rasztovits E, Cseke I, Bereczki K, Németh TM (2021) Different drought sensitivity traits of young sessile oak (Quercus petraea (Matt.) Liebl.) and Turkey oak (Quercus cerris L.) stands along a precipitation gradient in Hungary. Forest Ecology and Management 492: 119165.
Mölder A, Sennhenn-Reulen H, Fischer C, Rumpf H, Schönfelder E, Stockmann J, Nagel RV (2019) Success factors for high-quality oak forest (Quercus robur, Quercus petraea) regeneration. Forest Ecosystems 6: 1-17.
Nar H, Saglam A, Terzi R, Varkonyi Z, Kadioglu A (2009) Leaf rolling and photosystem II efficiency in Ctenanthe Setosa exposed to drought stress. Photosynthetica 47(3): 429-436.
Nyamjav B (2022) Adaptability responses to drought stress in the oak species Quercus petraea growing on dry sites. Journal of Forest Science 68(11): 459-472.
O'Brien E (1998) Water‐energy dynamics, climate, and prediction of woody plant species richness: an interim general model. Journal of Biogeography 25(2): 379-398.
Osakabe Y, Osakabe K, Shinozaki K, Tran LSP (2014) Response of plants to water stress. Frontiers in plant science 5: 86.
Öztürk S (2013) Turkey meşeleri teşhis ve tanı kılavuzu, Orman ve Su İşleri Bakanlığı, Orman Genel Müdürlüğü, Orman Zararlılarıyla Mücadele Dairesi Başkanlığı,Turkey.
Pirasteh‐Anosheh H, Saed‐Moucheshi A, Pakniyat H, Pessarakli M (2016) Stomatal responses to drought stress. Water stress and crop plants: A sustainable approach 1: 24-40.
Popović Z, Milanović S, Miletić Z, Smiljanić M (2010) Photosynthetic efficiency of Pedunculate oak seedlings under simulated water stress. Bulletin of the Faculty of Forestry 101: 139-150.
Pukacki PM, Kamińska-Rożek E (2005) Effect of drought stress on chlorophyll a fluorescence and electrical admittance of shoots in Norway spruce seedlings. Trees 19: 539-544.
Ranjan A, Sinha R, Singla‐Pareek SL, Pareek A, Singh AK (2022) Shaping the root system architecture in plants for adaptation to drought stress. Physiologia plantarum 174(2): e13651.
Ritchie GA, Shula RG (1984) Seasonal changes of tissue-water relations in shoots and root systems of douglas-fir seedlings. Forest Science 30(2): 538-548.
Ritchie GA (1984) Assessing seedling quality. Forest Nursery Manual: Production of Bareroot Seedling (ed. by ML Duryea and TD Landis) Martinus Nijhoff/Dr. W. Junk. Publishers, Hague/Boston/Lancaster.
Sankar B, Jaleel CA, Manivannan P, Kishorekumar A, Somasundaram R, Panneerselvam R (2007) Drought-induced biochemical modifications and proline metabolism in Abelmoschus esculentus (L.) Moench. Acta Botanica Croatica 66(1): 43-56.
Scholz FG, Bucci SJ, Arias N, Meinzer FC, Goldstein G (2012) Osmotic and elastic adjustments in cold desert shrubs differing in rooting depth: coping with drought and subzero temperatures. Oecologia 170: 885-897.
Semerci A (2002) Sedir (Cedrus libani A. Rich.) fidanlarına ait bazı morfolojik ve fizyolojik karakteristikler ile İç Anadolu’daki dikim başarısı arasındaki ilişkiler. İç Anadolu Ormancılık Araştırma Müdürlüğü Teknik Bülten No: 279, Ankara.
Shvaleva AL, Silva FCE, Breia E, Jouve J, Hausman JF, Almeida M, Maroco JP, Rodrigues ML, Pereıra JS, Chaves MM (2006) Metabolic responses to water deficit in two Eucalyptus globulus clones with contrasting drought sensitivity. Tree Physiology 26(2): 239-248.
Sircelj H, Tausz M, Grill D, Batic F (2005) Biochemical responses in leaves of two apple tree genotypes subjected to progressing drought. J. Plant Physiol. 162: 1308-1318.
Terzi R, Sağlam A, Kutlu N, Nar H, Kadioğlu A (2010) Impact of soil drought stress on photochemical efficiency of photosystem II and antioxidant enzyme activities of Phaseolus vulgaris cultivars. Turkish Journal of Botany 34(1): 1-10.
Thomas FM, Gausling T (2000) Morphological and physiological responses of oak seedlings (Quercus petraea and Quercus robur) to moderate drought. Annals of Forest Science 57(4): 325-333.
Tilki F (2010) Influence of acorn size and storage duration on moisture content, germination and survival of Quercus petraea (Mattuschka). Journal of Environmental Biology 31: 325-328
Turan ES (2018) Turkey'nin iklim değişikliğine bağlı kuraklık durumu. Doğal Afetler Çevre Dergisi 4(1): 63-69.
Turcsán A, Steppe K, Sárközi E, Erdélyi É, Missoorten M, Mees G, Mijnsbrugge KV (2016) Early summer drought stress during the first growing year stimulates extra shoot growth in oak seedlings (Quercus petraea). Frontiers in Plant Science 7: 193.
Tuttle CL, South DB, Golden MS, Meldahl RS (1987) Relationship between initial seedling height and survival and growth of loblolly pine seedlings planted during a droughty year. Southern Journal of Applied Forestry 11(3): 139-143.
Vernay A, Malagoli P, Fernandez M, Perot T, Améglio T, Balandier P (2018) Improved Deschampsia cespitosa growth by nitrogen fertilization jeopardizes Quercus petraea regeneration through intensification of competition. Basic and Applied Ecology 31: 21-32.
Wright A, Schnitzer SA, Reich PB (2015) Daily environmental conditions determine the competition-facilitation balance for plant water status. Journal of Ecology 103: 648-656.
Wu M, Zhang WH, Ma C, Zhou JY (2013) Changes in morphological, physiological and biochemical responses to different levels of drought stress in chinese cork oak (Quercus variabilis BI.) seedlings. Russ. J. Plant Physl. 60(5): 681-692.
Xiong S, Wang Y, Chen Y, Gao M, Zhao Y, Wu L (2022) Effects of drought stress and rehydration on physiological and biochemical properties of four oak species in China. Plants 11(5): 679.
Yahyaoğlu Z, Genç M (2007) Fidan standardizasyonu. Standart fidan yetiştirmenin biyolojik ve teknik esasları, SDÜ Orman Fakültesi Yayın No: 75, Isparta, Turkey.
Yang Y, Liu Q, Han C, Qiao YZ, Yao XQ, Yin HJ (2007) Influence of water stress and low irradian on morphological and physiological characteristics of Picea asperata Masters. seedlings, Photosynthetıca 45(4): 613-619.
Yavaş İ, Akgül HN, Ünay A (2016) Bitkilerin kuraklığa dayanıklılığını arttırmaya yönelik uygulamalar. Türk Tarım-Gıda Bilim ve Teknoloji Dergisi 4(1): 48-57.
Zolfaghari R, Akbarinia M (2018) Physiological and biochemical responses of Quercus brantii seedlings to water deficit stress. Caspian Journal of Environmental Sciences 16: 165-177.



