Schlüsselbegriffe: Pappeln, sexuelle Vermehrung, Vermehrung durch Stecklinge
Available at https://doi.org/10.53203/fs.2401.1
See below the issue 1/2024 as E-Paper or have a look at our E-Paper archive dating back to 1955.
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Abstract
To produce seed-originated seedlings of Populus caspica Bornm. and compare them with mother trees, mature capsules were collected from 17 native Caspian poplar trees from the three provinces of Gilan, Mazandaran, and Golestan in northern Iran. After the successful mass production of seedlings from these 17 mother trees, three half-sib offspring of each of these 17 mother trees and the seedlings obtained from cuttings of the mother trees were selected and planted in a randomized complete block design with three replications. 13 leaf morphological traits, collar diameter and survival rate were measured at the end of the growing season, and the plant height was measured in six one-month periods. Results showed a significant difference in height growth between the mother trees and offspring in all months except August and October. The offspring growth rate during all of the periods was significantly higher than the growth rate of the cuttings from the mother trees. The mean height growth of the offspring and mother trees were 285.1 and 250.4 cm, respectively, as well as the mean diameter growth of the offspring and mother trees were 20.9 and 18.2 mm respectively. Also, there were significant differences in the height growth rate between the origin (mother trees), times, offspring, interaction effects of offspring*time, and offspring*origin. Results showed vast variations between the offspring in several morphological traits that could be used to produce new varieties of the species. Further investigating the offspring data revealed that 13 traits showed significant differences, while in mother tree data only six traits showed significant differences between the mother trees. The average of 15 traits measured in the offspring was higher than for the mother trees and only the mean of the three traits in the offspring was lower than for the mother trees. Plant diameter and height showed a significant positive correlation to leaf size traits. Plant diameter and height showed a significant positive correlation to leaf size traits. These traits were higher in the offspring compared to the mother trees. Significant differences between the offspring and mother trees, especially the superiority of most of the offspring traits, indicated an increase in genetic diversity and improvement in the growth of the offspring derived from the sexual reproduction of this precious Hyrcanian species.
Zusammenfassung
Reifes Saatgut von Populus caspica Bornm. wurden von 17 Bäumen in den Provinzen Gilan, Masandaran und Golestan im Norden vom Iran gesammelt, um Sämlinge zu produzieren und mit den Mutterbäumen zu vergleichen. Nach der erfolgreichen Massenproduktion der Sämlinge dieser 17 Mutterbäume wurden drei Halbgeschwister-Sämlinge zusammen mit aus Stecklingen von den Mutterbäumen erhaltenen Sämlingen in Form eines randomisierten vollständigen Blockdesigns mit drei Wiederholungen gepflanzt. Am Ende der Vegetationsperiode wurden 13 Blattmorphologiemerkmale, der Stammdurchmesser und die Überlebensrate sowie die Pflanzenhöhe in sechs einmonatigen Zeiträumen gemessen. Die Ergebnisse zeigten einen signifikanten Unterschied im Höhenwachstum zwischen den Mutterbäumen und den Nachkommen in allen Monaten außer August und Oktober. Die Wachstumsrate der Nachkommen war in allen Zeiträumen deutlich höher als die ihrer Mutterbäume. Das mittlere Höhenwachstum der Nachkommen- und Mutterbäume betrug 285,1 cm bzw. 250,4 cm und das mittlere Durchmesserwachstum der Nachkommen- und Mutterbäume betrug 20,9 mm bzw. 18,2 mm. Außerdem gab es signifikante Unterschiede in der Höhenwachstumsrate zwischen Herkunft (Mutterbäume), Zeiten, Nachkommen, Interaktionseffekten von Nachkommen*Zeit und Nachkommen*Herkunft. Die Ergebnisse zeigten, dass es eine große Vielfalt in manchen morphologischen Merkmalen der Nachkommen gibt, die zur Produktion neuer Varianten zu nutzen sein können. Bei der Untersuchung der Daten von Nachkommen wurden 13 Merkmale gefunden, die signifikante Unterschiede verursachten, während in den Daten der Mutterbäume nur 6 Merkmale signifikante Unterschiede bei den Mutterbäumen zeigten. Durchschnittlich waren 15 Merkmale bei den Nachkommen mehr als bei den Mutterbäumen und nur durchschnittlich 3 Merkmale bei den Nachkommen waren weniger als bei den Mutterbäumen. In der Korrelationsanalyse wurde eine deutliche Korrelation zwischen den Merkmalen des Durchmessers und der Höhe und Merkmale der Blattabmessungen festgestellt. Die signifikanten Unterschiede zwischen den aus der sexuellen Vermehrung hervorgegangenen Sämlingen und den Stecklingen der Mutterbäume deuten auf die Zunahme der genetischen Vielfalt der aus der sexuellen Vermehrung hervorgegangenen Sämlinge dieser wertvollen Art der Hyrkanischen Wälder hin.
1 Introduction
Populus caspica Bornm. (Caspian poplar) was first studied by Bornmuller (Bornmuller, 1908), which according to The WFO Plant List (https://wfoplantlist.org/plant-list/) has a synonym with Populus alba var. caspica Bornm. In some sources, it is called Populus alba L. (Sabeti, 1976). Because of the many morphological similarities between P. caspica Bornm. and P. alba, some botanists argue that these two species, belong to the same species (Rechinger, 1982). Several molecular markers have been applied to differentiate the species (Khasa et al., 2003; Cervera et al., 2004; Wan et al., 2013). P. caspica Bornm. has very similar morphological features with P. alba, but is differentiated mainly by the inflorescence and petiole sizes; the petiole of P. caspica Bornm. is 10-15 cm long (P. alba 10 cm) and inflorescence is 20 cm long (P. alba 10-15 cm). P. alba has many ridges in its bark and grows in the sun, tolerates almost any soils, wet or dry, and is distributed in the river areas of southern Europe, north Africa and central Asia; this species can be found in active floodplains (Hamzeh and Dayanandan, 2004). P. caspica Bornm. with few ridges in its bark (Rechinger, 1982), is distributed exclusively in the Hyrcanian Forest with other relict tree species, such as Alnus glutinosa, Pterocarya fraxinifolia, and Quercus castaneifolia (Yousefzadeh et al., 2019). Caspian poplar has a high tolerance to periodic flooding, which is typical for its natural habitat (Sadati et al., 2010). The habitat of this endemic poplar species has undergone genetic erosion due to uncontrolled logging (Gamari Zare et al., 2020). This valuable species is endangered according to the IUCN red list (Fallahchai et al., 2016). So, it is now designated as a protected species by the Environmental Protection Agency of Iran (Jafari Mofidabadi, 2005). The primary threat to the native poplar species, such as Caspian poplar, is human encroachment, particularly agricultural development, residential development, and sand mining in riverbeds (Alipour et al., 2021a).
This species requires an open bed, fertile soil, and sufficient moisture, not necessarily flooding, to regenerate naturally. Although there used to be large and pure stands of this species in northern Iran, its number, distribution, and regeneration have declined due to various factors (Jalili and Jamzad, 1999), including climate change and habitat conditions. Sunlight, suitable soil, and sufficient moisture are necessary for the successful germination of poplar seeds. But they are often not realized in natural conditions (Asadi and Mirzaie Nodoushan, 2011). Occasionally Caspian poplar seedlings are observed in some areas of Hyrcanian Forests, but studies have shown that they are often the result of regeneration from coppice, not from seed and due to the low rate of sexual reproduction in nature, these trees have insufficient genetic diversity (Asadi, 2015). With the help of sexual reproduction and increasing genetic diversity, both the possibility of mass production of seedlings and the expansion of the genetic base of these trees can be provided, and as a result, among the new genotypes can be found clones with a comparative advantage over the conventional clones. The new clones will be an ideal option for the reforestation of deforested areas in the north of Iran. Poplar habitat degradation has led to increasing geographic distances between populations and fragmentation of their natural habitats. Fragmentation has negatively affected gene flow (Alipour et al., 2021b; Young et al., 1996; Dayanandan et al., 1999). In addition, this phenomenon has reduced the survival rate in large populations (Hanski and Ovaskainen, 2000; Casagrandi and Gatto, 2002), increased consanguinity within populations, and greater susceptibility to perturbations or demographic stochasticity (Imbert and Lefevre, 2003). In most riparian species, seeds are carried by the wind and transported through the water. Seeds are thought to cover several kilometres and gene flow is therefore supposed to be continuous and to give rise to large population size. Furthermore, the direction of dispersal is expected to be predominantly downstream. Thus, an accumulation of genetic diversity in populations located in the lower reaches of a river can be expected (Imbert and Lefevre, 2003). Alipour et al. (2021b), to understand how anthropogenic and environmental factors have influenced the pattern of genetic diversity of P. caspica did research based on 14 microsatellite markers. Nine populations showed a significant reduction in effective population size and a genetic bottleneck. To determine ecotypes of P. caspica in plain communities of Mazandaran and Gilan provinces, Fallah et al. (2011) measured 13 morphological traits in addition to enzymatic analysis and based on cluster analysis, indicated three different ecotypes. Based on research done on other Poplar species Alimohammadi et al. (2012) found the low genetic variation in P. nigra stands in the Kermanshah province of Iran due to asexual reproduction. They recommended providing conditions for sexual reproduction and planting seedlings of different clones to increase the genetic richness of the existing populations. Similarly, Homaie et al. (2014) investigated several morphological and micro-morphological traits on four half-sib progenies of each eleven parental clones of P. euphratica stands in the Karaj province of Iran. Results indicated a vast variation between the studied parents and their progenies that may be used to produce new commercial varieties of the species. Yu and Tigerstedt (2001) studied four 5-year-old hybrid clones (P. tremula × P. tremuloides) and one local clone, P. tremula seedling source, based on some growth traits and observed differences in phenological and growth traits among hybrid clones and P. tremula. Caspian poplar trees have optimal distribution in sunny and warm valleys. Global temperatures are rising (Abbass et al., 2022), so species with relative heat tolerance will receive special attention in the future as suitable afforestation options in warmer regions. Asadi and Mirzaie Nodoushan (2011) mentioned the reduction in poplar genetic diversity due to asexual propagation was presented as a reason for conducting breeding activities and using their seeds for propagation. In suitable microhabitats, many seeds originated seedlings are established (sometimes up to 4000 seedlings per m2), but the mortality rate is also very high (more than 77 to 100%), and persistent waterlogging may be responsible for the mortality of newly germinated and older seedlings (Dixon and Turnel, 2006). In addition, Fang (2012) compared the volumetric growth of 5-year-old trees of P. alba and P. alba L. × P. glandulosa hybrids. The results showed that the hybrids exhibited greater root growth and produced 30% more wood than their parents. Benetka et al. (2002) also compared the production capacity of P. nigra with three hybrids by measuring the diameter and height growth. They introduced suitable clones for cultivation on marginal agricultural land.
Using various traits in the form of experimental design, this study attempted to determine the difference in vegetative characteristics of Caspian poplar offspring and their mother trees grown in the three provinces of Gilan, Mazandaran, and Golestan in northern Iran. The aims of present investigation are: (1) to assess the growth performance of half-sib offspring and mother trees based on height, diameter growth and survival (2) to estimate the genetic diversity on the basis of morphological traits as easy and cheap marker to find out about genetic diversity initially (Alimohamadi et al.,2012), and (3) to compare the function of different generations based on different growth and morphological traits.
2 Materials and methods
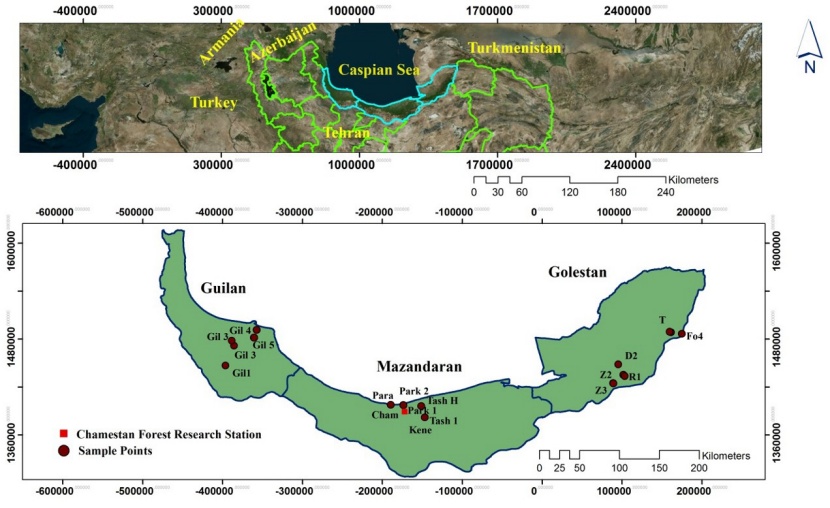
Figure 1: Location of Gilan, Mazandaran and Golestan provinces in the north of Iran (up) and position of samples in the three provinces (down).
Abbildung 1: Geografische Lage der Provinzen Gilan, Masandaran und Golestan im Norden vom Iran (oben) und die Lage der Proben in den drei Provinzen (unten).
2.1 Sampling and experiment design
Initially, 17 native Caspian poplar trees from the provinces of Gilan, Mazandaran, and Golestan in northern Iran (Figure 1) were selected (location and main characteristics of sampled trees habitat and trees are shown in Table 1). Then the opening capsules were collected from each tree in April. All the capsules were kept completely isolated to complete their flowering stages. A large number of seeds were separated from the capsules of each tree and planted after complete sifting and cleaning (due to the absence of introduced species in the region, seeds are not hybrids). After the successful mass production of seedlings belonging to 17 mother trees, the cuttings of the three best (in terms of dimensions and freshness) seed originated seedlings from each mother tree and the cuttings of the mother tree (totally 68 treatments including four samples of each 17 trees) were selected and planted in Chamestan Forest Research Station in Noor city (36° 25' N & 51° 55' E, 70 m a.s.l), Mazandaranprovince of Iran. Mean annual temperature and precipitation are 15.8 °C and 840 mm, respectively. The absolute minimum and maximum monthlyntemperatures are -8.5 °C and 36 °C, respectively. The average relative humidity is 78%. After one year of growing and producing uniform cuttings, 15 cuttings (five observations per three replications) with the same dimensions and freshness of each 68 treatments were planted in a randomized complete block design with three replications (totally 1020 cuttings). All maintenance activities, including irrigation and weeding, were carried out at scheduled times during the growing season.
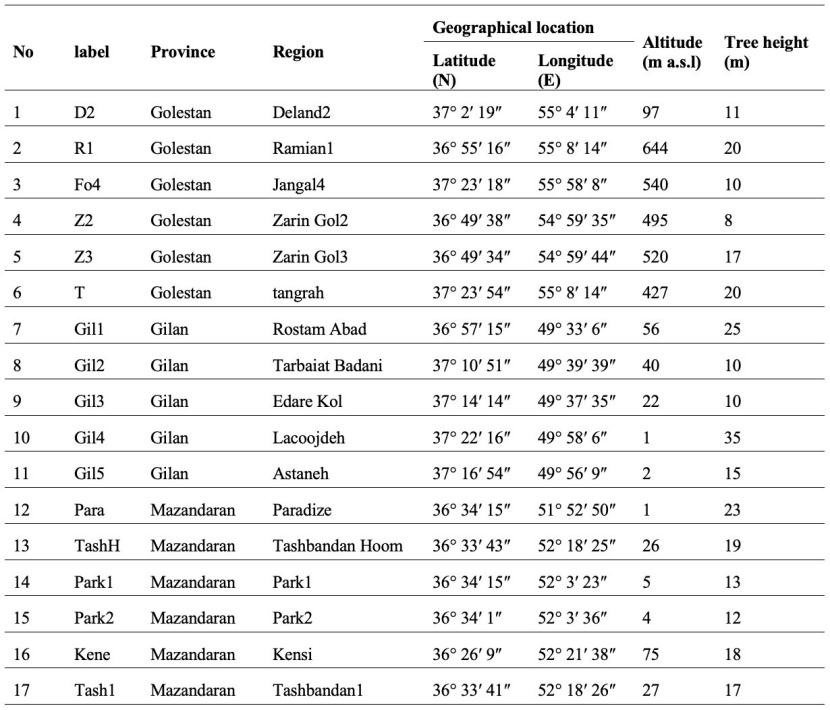
Table 1: Location and main characteristics of sampled trees habitat.
Tabelle 1: Geografische Lage und die wichtigsten Merkmale der beprobten Bäume.
2.2 Measured objects and statistical analyses
The difference between the mother trees and their progenies was determined by examining 18 traits. In addition to the morphological characteristics (Alba et al., 2002) listed in Table 2, collar diameter (in mm) and survival rate (in percent) were measured at the end of the growing season, and the height (in cm) was also measured in six one-month periods. Kolmogorov-Smirnov test for assessing the normality of the data, Levene test for determining the homogeneity of variance, independent t-test for comparing the mean characteristics of the mother trees and offspring, mixed ANOVA for comparing plant diameter and height at different periods, and principal component analysis for visualizing and categorizing the mother trees data in a more interpretable way (Alimohammadi et al., 2009) were conducted using SPSS 20. Because the offspring data were collected in a nested form, the nested model with the GLM option of SAS software was used (Homaie et al., 2014) to determine the differences in 15 morphological traits between the offspring and mother trees. Grouping has been done by the Duncan method. Pearson correlation coefficient between all dual compounds of the reported traits from the offspring was also calculated using SPSS 20 to investigate the relationship between these traits.
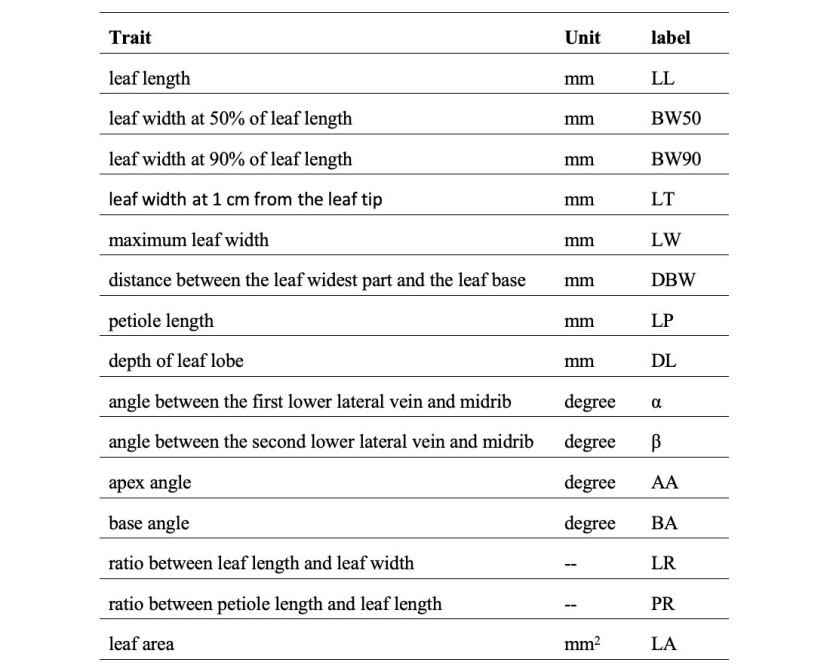
Table 2: Names, units and abbreviations of the studied morphological traits.
Tabelle 2: Namen, Einheiten und Abkürzungen der morphologischen Merkmale.
3 Results
The Kolmogorov-Smirnov test showed data had been drawn from a normally distributed population. The comparison of the mother trees and offspring performance based on the values of different traits is shown in Table 3. What is remarkable about these differences is the range of variation observed in the mother trees and their progenies. For all traits, the range of variation in the offspring was more than in mother trees except for six traits: BW50, LT, β, LR, PR, and diameter. For example, the range of variation for LL in the parent's leaves was 85 mm, whereas in the offspring was 120 mm. In other words, a new diversity was created in the progenies, which is important. The average of most of the studied traits increased significantly in offspring. Of 18 traits, 13 showed significant differences between the offspring and mother trees (there was no significant difference in BW50, DBW, β, AA, and PR). Of these traits, only LT and LR in the mother trees were larger than the offspring, while in the remaining 11 traits, especially diameter and height, the offspring were superior.
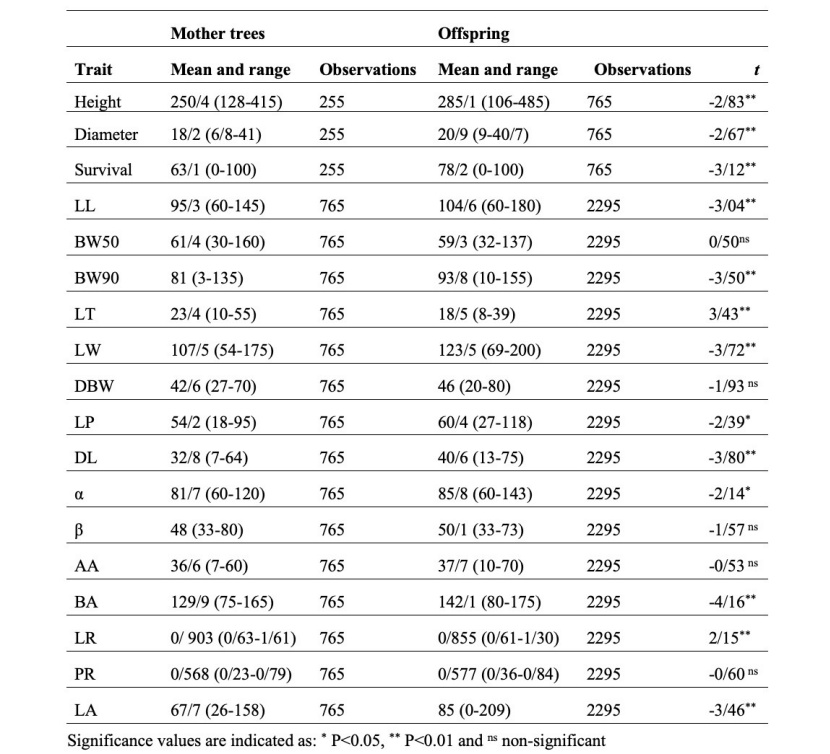
Table 3: Observations, mean values and ranges of variation (in brackets) of the studied traits and independent t-test for comparing the differences between the mother trees and offspring.
Tabeklle 3: Beobachtungen, Durchschnittswerte und Variationsbreite (in Klammern) der untersuchten Merkmale und unabhängige t-Test-Statistiken zum Vergleich des Unterschieds zwischen Mutter- und Nachkommenbäumen.
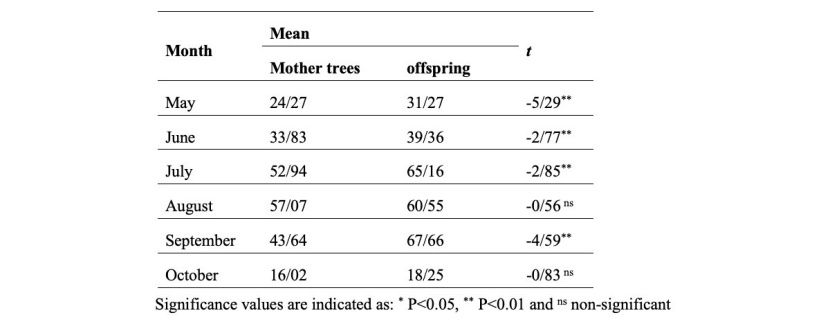
Table 4: Independent t-tests for comparing the differences between the mother trees and offspring in terms of height growth in the studied months.
Tabelle 4: Unabhängigs t-Tests zum Vergleichen des Unterschieds zwischen Mutter- und Nachkommenbäumen hinsichtlich des Höhenzuwachses in Untersuchungsperiode.
Independent t-test results (Table 4) showed a significant difference in height growth between the mother trees and offspring in all months except August and October. In addition, the growth rate of the offspring was always greater than the mother trees (Figure 2).
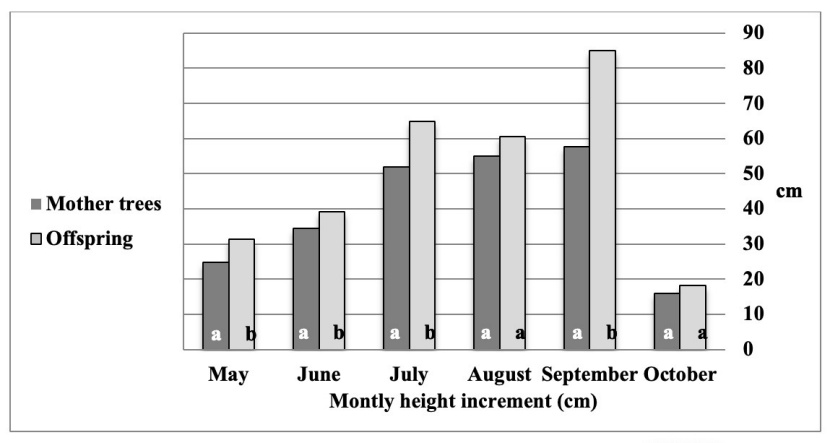
Figure 2: Comparison of mean height growth of the mother trees and offspring in different times according to t-test (similar letters indicate the same growth).
Abbildung 2: Vergleich des durchschnittlichen Höhenzuwachses von Mutterbäumen und Kindern in verschiedenen Zeiträumen gemäß t-Test (ahnliche Buchstaben weisen auf dasselbe Wachstum hin).
ANOVA results (Table 5) showed significant differences in the height growth rate between the origin (mother trees), times, offspring, interaction effects of offspring*time, and offspring*origin.
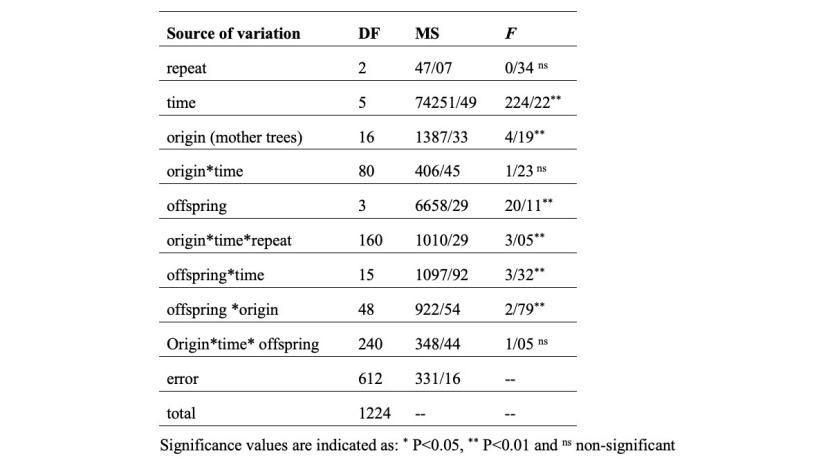
Table 5: Summary of the ANOVA of height growth.
Tabelle 5: Zusammenfassung der Ergebnisse der Varianzanalyse des Höhenzuwachses.
Results of nested ANOVA for height and diameter of total samples (the mother trees and offspring) (Table 6) revealed no significant difference between the mother trees based on these traits. ANOVA results of diameter, height, and survival rate of offspring showed no significant differences between the mother trees (Table 6). Still, the effect of offspring nesting in mother trees was significant only for survival rate. Also, ANOVA results of leaf morphological traits of total samples (Table 7) indicated significant differences between mean values of LL, DBW, LP, and PR traits. The effect of offspring nesting in mother trees was significant in α, β, and LR traits. According to Table 7, which presents the results of ANOVA for leaf morphological traits of the offspring, the mother trees differed significantly in the leaf characteristics of their offspring based on LL, BW90, maximum leaf width (LW), DBW, LP, and LA traits. The effect of offspring nesting in mother trees was significant only for the β trait.
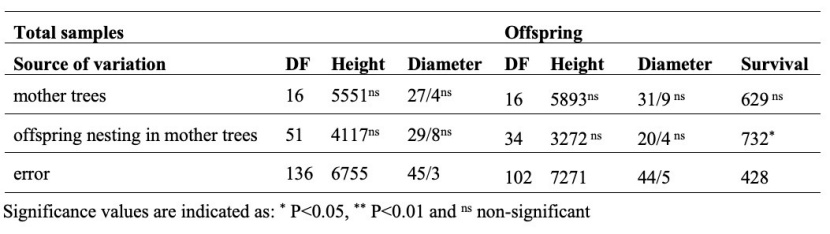
Table 6: Summary of the nested ANOVA of height and diameter of total samples and height, diameter and survival of offspring.
Tabelle 6: Zusammenfassung der Ergebnisse der Varianzanalyse von nested-factor-Modell von Höhe und Durchmesser und von Höhe, Durchmesser und Überlebensfähigkeit von Nachkommen (Kinder).
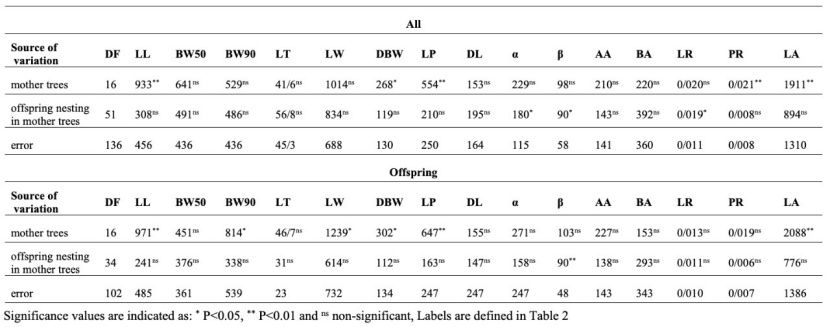
Table 7: Summary of the nested ANOVA of leaf morphological traits of all samples and only offspring samples.
Tabelle 7: Zusammenfassung der Ergebnisse der Varianzanalyse von nested-factor-Modell von morphologischen Blattmerkmalen von allen Proben und nur von Nachkommen (Kinder).
Mother trees grouping based on the mean of the offspring and parental traits (total) by the Duncan method, LW, BA, LA, DL, BW90, LT, diameter, α, and β traits were in the same group. In contrast, the other traits (nine traits) listed in Table 8 were classified into different groups. Also, according to mother trees, grouping based on the mean of the offspring traits (Table 9) BA, DL, diameter, and height traits were classified in the same group, while the other traits listed in Table 9 (13 traits) were arranged in separate groups.
Mother trees grouping based on the mean of the parental traits by the Duncan method (Table 10) revealed no significant differences in the grouping of mother trees based on LL, BW90, LT, LW, DBW, LP, BA, and LA traits. They were all in the same group but based on the other characteristics listed in Table 10 (seven characteristics), the mother trees were categorized in separate groups. Based on Table 10, most traits (LL, BW90, LT, LW, DBW, LP, BA, and LA) did not show significant differences in parental separation, and a high level of similarities occurred between the mother trees. However, according to Table 9, only the two morphological traits, BA and DL, did not show significant differences in offspring separation. Therefore, the diversity of offspring was very remarkable. Moreover, when mother trees were grouped according to the offspring traits, more groups emerged, indicating greater diversity among the offspring.
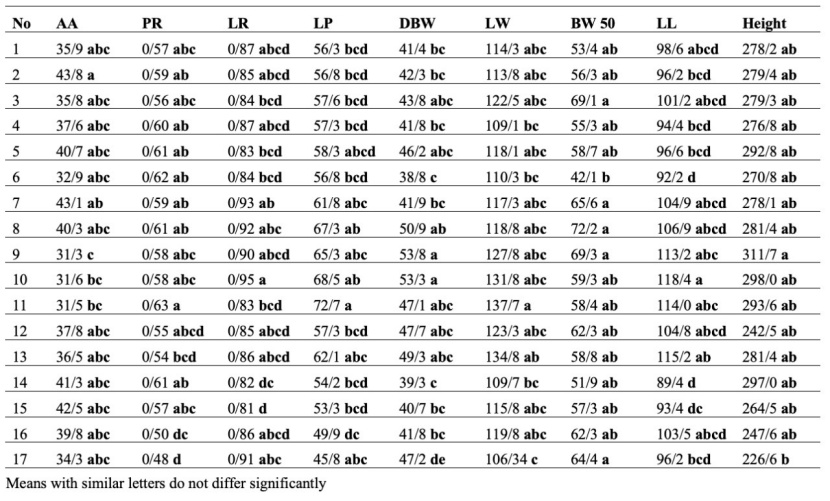
Table 8: Mother trees grouping based on the mean of the offspring and parental traits (total) by Duncan method.
Tabelle 8: Gruppierung der Mutterbäume nach Durchschnittsmerkmalen der Kinder und Eltern (gesamt) durch Duncan Prozedur.
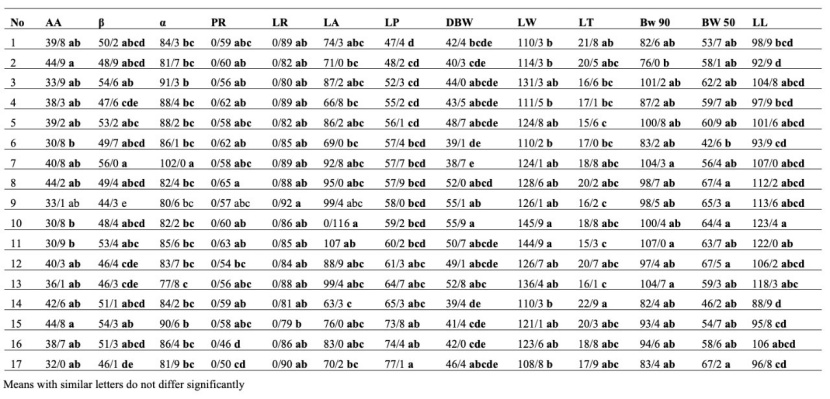
Table 9: Mother trees grouping based on the mean of the offspring traits by Duncan method.
Tabelle 9: Gruppierung der Mutterbäume nach dem Mittelwert der Merkmale der Kinder mittels Duncan Prozedur.
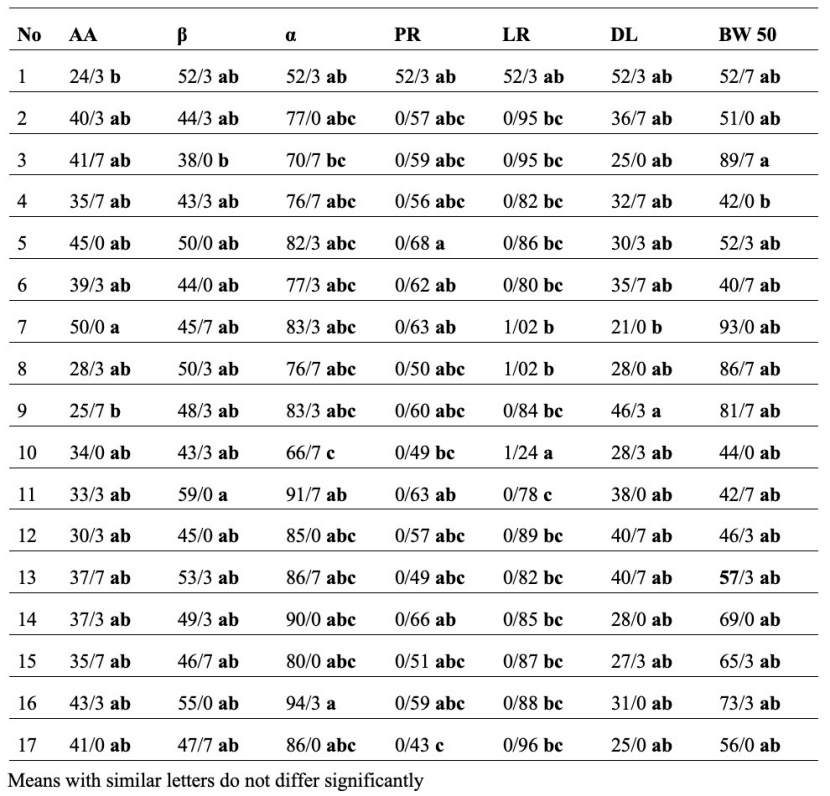
Table 10: Mother trees grouping based on the mean of the parental traits by Duncan method.
Tabelle 10: Gruppierung der Mutterbäume nach dem Mittelwert der elterlichen Merkmalen mittels Duncan Prozedur.
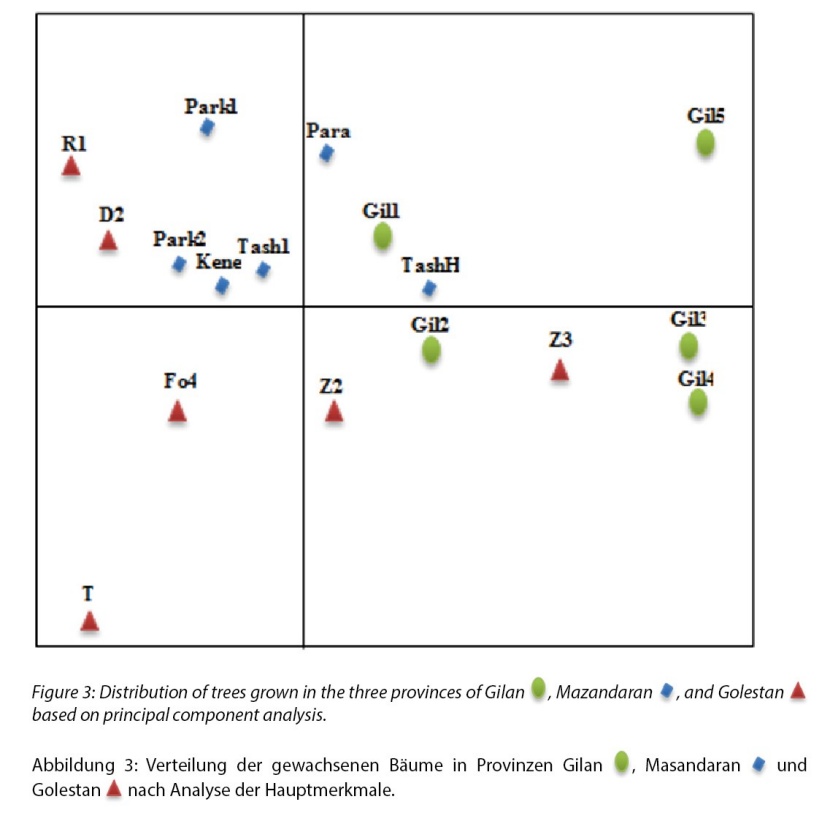
Figure 3: Distribution of trees grown in the three provinces of Gilan, Mazandaran, and Golestan based on principal component analysis.
Abbildung 3: Verteilung der gewachsenen Bäume in Provinzen Gilan, Masandaran und Golestan nach Analyse der Hauptmerkmale.
Based on the principal component analysis, 10 components were obtained, which covers 98.4% of the variance in total. The first, second and third principal components comprise 40.49%, 16.72%, and 12.49% of the total variance respectively, and in total the first three principal components comprise 70% and the first two principal components comprise 57% of the total variance. The results of the principal component analysis (Figure 3) showed the distribution of the different mother trees on the coordinate axis using the first and second principal components in such a way that the values of the most effective traits increased from left to right in the first component and from bottom to top in the second component. Therefore, the mother trees with the highest value in the first principal component (Gilan province samples) were those with greater amounts in LA, LW, LL, and DL traits. So, the trees from Gilan province are situated on the right side of the first component with greater amounts, but the tree Z3, which belongs to Golestan province, was also in this part. Correlation test results (Table 11) showed that the LA trait was significantly correlated with more traits (10 out of 16 traits). Height was significantly associated with the diameter, and these two traits had a significant correlation with traits related to leaf dimensions, including LL, BW50, BW90, LT, LW, DBW, LP, DL, β, PR, and LA traits. Also, the α trait was correlated with only four traits: DBW, β, BA, and LR. However, the β trait is significantly associated with ten traits: height, diameter, BW50, LT, LW, DBW, DL, α, BA, and LR. Among the studied traits, the LW trait had the highest number of correlations, so that, except for the α trait, it had a significant correlation with other traits.
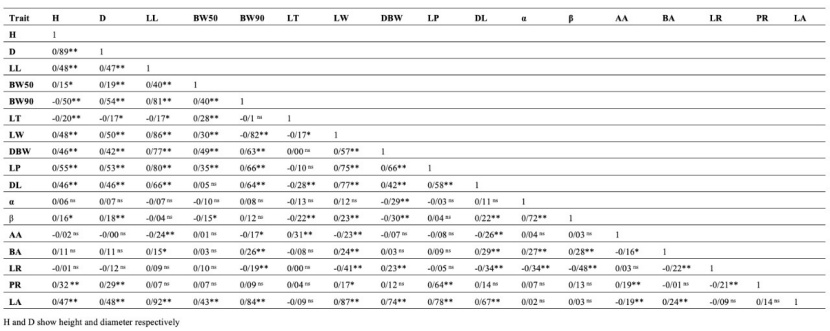
Table 11: Correlation coefficients between the studied traits and their level of significance.
Tabelle 11: Korrelationskoeffizienten zwischen untersuchten Merkmalen und ihrem Signifikanzniveau.
4 Discussion
Examination of the height growth of all seedlings in one-month periods of the growing season, the significant difference in time, origin, and their interactions between and within the origin and offspring indicates the existence of high variation in height growth between them. The three months with the highest percentage growth of all seedlings were July, August, and September. An important aspect of the study is the high growth rate in September, which is different from the results of Alimmohammadi et al. (2015), conducted for P. alba and P. nigra in Alborz province, while similar to the results of Yousefi and Modirrahmati (2004) shown in Kurdistan province. Alimohammadi et al. (2015) reported the highest growth in July and the lowest in September for P. nigra. Yousefi and Modir Rahmati (2004), on the other hand, identified three different vegetation stages, including the initial slow growth stage (before June), the rapid growth stage (June to September), and the secondary slow growth stage (after September). The differences between the studied species and areas effectively affected these different results. Therefore, phenological studies should be planned separately for each geographical location and species.
Nevertheless, the study of the vegetative behavior of species is essential to determine the potential growth of trees under different environmental conditions, as some species have now been shown to have more significant growth potential than previously believed (Christerson, 2006). In Table 4, it is shown that the height growth of offspring was significantly greater than the mother trees in 4 months but was not significantly different in the remaining two months. The superiority of offspring over mother trees in both monthly and final growth confirms the need to pay more attention to sexual reproduction in these trees. Little-observed natural regeneration of P. caspica is due to asexual reproduction (coppics), and sexual regeneration is rare in the habitats of these trees. Alipour et al. (2021b) revealed that the Alborz mountains are a major barrier to gene flow between P. caspica populations in the Hyrcanian forests. Poplar seeds need sufficient moisture, light, and fertile soil to continue growing, and these three agents are rarely present simultaneously under the natural conditions of their habitats. Therefore, to develop sexual reproduction of these trees in the nurseries, it is necessary to provide suitable conditions in terms of the three primary factors of light, moisture, and soil, so that relying on the achievements of the present study, which has shown the superiority of offspring over mother trees it may be possible to restore the deforested areas in the north of Iran by planting P. caspica trees. Therefore, more attention should be paid to the sexual reproduction of these trees.
Based on the results of ANOVA for total samples (both mother trees and offspring) (Table 7), there were significant differences between the mother trees in terms of LL, DBW, LP, LA, and PR traits which all related to leaf dimensions. Still, there were no significant differences between the mother trees for other traits. However, nested analysis of the mother trees revealed a significant difference between only three traits: α, β, and LR, and there were no differences in all other characteristics. However, according to Table 7, which is based solely on the offspring data, the BW90 trait also showed significant differences in addition to the above traits. It is noteworthy that in these data for the effect of offspring nesting in the mother trees, just two traits, β and survival rate, differed significantly. According to these results, it can be concluded that, firstly, in most of the 18 traits, no significant differences were found between the mother trees (despite being located in different provinces); secondly, the offspring of different parents did not show significant differences except for two traits related to leaf angle. However, a grouping of the mother trees based on Duncan's test revealed more groups among the offspring than the mother trees. Thus, the diversity of the offspring was greater than that of the mother trees (Tables 8 and 9). Mean values of offspring diameter and height showed relative superiority over the mother trees.
The observed diversity in the mother trees and offspring was generally lower than expected. This situation may be explained by the destruction of the natural habitats of this species in the north of Iran and the absence of sexual reproduction in the past. Notably, although pollination of the trees in the present study occurred naturally, it is possible; it had been done between neighboring trees. Therefore, similarities between the mother trees and their progenies are expected. If artificial inbreeding occurs between distant parents, greater diversity will be possible.
The results of this study are in contrast to Homaie et al. (2014), who observed a large variation between the parents and progenies of P. euphratica but in our findings P. caspica showed a lower variation. A key aspect of their research is that parental trees with various origins from different provinces were pollinated and outbred at the Alborz Research Station in Karaj province. The progenies of parents with different origins are more diverse.
The range of variation of 12 traits was higher in the offspring than in the mother trees. This conclusion is comparable with the work of Homaie et al. (2014), which showed vast variation between parents and their progenies based on the studied traits. Increasing the range of traits is of particular importance. By increasing the range of traits, it can be expected that new capacities can be obtained that can be used in breeding programs to produce new commercial varieties of the specie (Homaie et al., 2014).
By analyzing the principal components and determining the traits most important in separating and grouping the mother trees, the situation of Mazandaran samples located between Gilan and Golestan samples was a noteworthy point. This situation can indicate the segregation based on geographical distribution because the Mazandaran province is between Gilan and Golestan. These results are in accordance with the results of Alimohammadi et al. (2009), which, based on analyzing the principal components, showed that P. nigra stands belonging to the two provinces were separated based on morphological traits.
Using correlation analysis, height and diameter, in addition to each other with traits related to leaf dimensions, including LA, showed the highest significant correlations. The same results have been indicated in some studies on poplar species (Asadi et al., 2005; Alimohamadi et al., 2015). There is considerable research showing that in the poplar genus, leaf area trait (through the effect on light absorption) is associated with production and can be used as an indicator for selecting trees with high production (Marron et al., 2008; Bunn et al., 2004; Rae et al., 2004).
5 Conclusion
The results of the morphological analysis showed that the offspring were more diverse than the mother trees in terms of the studied traits and the average of most traits measured in the offspring was higher than the mother trees as well as the growth rate of the offspring was significantly higher than the mother trees. Since the study of leaf morphological traits is one of the necessary measures in estimating genetic diversity, separation of ecotypes, and breeding activities of poplar (Storme et al., 2002; Lopez et al., 2004; Calagari et al., 2006) and according to the results of the present study, which indicate a lack of diversity between parental trees growing in natural, but often destroyed habitats, it is recommended that samples of these valuable and endangered species should be collected throughout the northern provinces of Iran and kept as a collection in a suitable field to conserve these heritage reserves and increase diversity in the gene bank. Phenological, morphological, and breeding studies should also be carried out on them. In addition, this collection allows sexual intercourse between distant trees to create more diversity.
References
Abbass K, Qasim MZ, Song H, Murshed M, Mahmood H, Younis I (2022). A review of the global climate change impacts, adaptation, and sustainable mitigation measures. Environmental Science and Pollution Research 29: 42539-42559. https://doi.org/10.1007/s11356-022-19718-6
Alba NC, Maestro D, gundez MD, Notivol E (2002). Advances in the preservation of genetic resources in Populus nigra L. in Spain. In: Van Dam BC, Bordacs S, (editors). Genetic diversity in river populations of European black poplar. Budapest, Hungary: Csiszar Nyomda Ltd, pp. 125-136.
Alimohammadi A, Asadi F, Tabaei Aghdaei R (2015). Evaluation of growth and morphological parameters in two poplar species (P. nigra l. & P. alba l.) to tree growth reveal traits related to productivity (case study in Kermanshah, Zanjan and Esfahan provinces). Ecology of Iranian Forests 3 (5): 31-41 (in Persian with an abstract in English). http://ifej.sanru.ac.ir/article-1-184-en.html
Alimohammadi A, Asadi F, Tabaei Aghdaei R (2012). Genetic diversity in Populus nigra plantations from west of Iran. Annals of Forest Research 56 (1): 165-178. https://www.afrjournal.org/index.php/afr/article/view/56
Alimohammadi A, Asadi F, Adeli E, Tabaei Aghdaei R, Mataji A (2009). Using morphological traits for identification of Populus nigra stands. Iranian Journal of Forest and Poplar Research 7 (3): 369-381 (in Persian with an abstract in English). https://ijfpr.areeo.ac.ir/article_107819.html?lang=en
Alipour S, Badehian Z, Yusefzadeh H, Espahbodi K, Asadi F (2021a). Predicting the suitable habitat of Caspian poplar (Populus caspica Bornm.) and the most influential climatic variables on its potential distribution in the Hyrcanian forests, Iran. Iranian Journal of Forest and Poplar Research 29 (2): 176-188 (in Persian with an abstract in English). https://doi.org/10.22092/IJFPR.2021.354716.2001
Alipour S, Yousefzadeh H, Badehian Z, Asadi F, Espahbodi K (2021b). Genetic diversity and structure of the endemic and critically endangered Populus caspica in the Hyrcanian forests. Tree Genetics and Genomes 17 (2): 1-13. https://doi.org/10.1007/s11295-021-01497-9
Asadi F (2015). Sexual regeneration and seed bourn seedling production for expanding genetic variation of Populus caspica Bornm. for introducing new clones, the final report of the research project. Agriculture and natural Resources Research Center of Mazandaran province. p 56 (in Persian).
Asadi F, Mirzaie Nodoushan H (2011). Evaluation of different treatments in sexual reproduction of Populus caspica Bornm. for broadening its genetic basis in the nature. Iranian Journal of Forest and Poplar Research 19 (3): 441-452 (in Persian). https://www.sid.ir/en/Journal/ViewPaper.aspx?ID=251591
Asadi F, Mirzaie Nodoushan H, Modir Rahmati A (2005). Path analysis of poplar different attributes in early stages of growth. Iranian Journal of Forest and Poplar Research 13 (3): 295-312 (in Persian). https://ijfpr.areeo.ac.ir/article_108500.html
Benetka V, Bartakova I, Mottl J (2002). Productivity of Populus nigra L. ssp. nigra under short-rotation culture in marginal areas. Biomass and Bioenergy 23 (5): 327-336. https://doi.org/10.1016/S0961-9534(02)00065-X
Bornmuller JFN (1908). In: Beuverd G (eds.), Bulletin de l'Herbier Boissier 2, p 724.
Bunn SM, Rae AM, Herbert CS, Raylor G (2004). Leaf- level productivity traits in Populus grown in short rotation coppice for biomass energy. Forestry 77 (4): 307-323. https://doi.org/10.1093/forestry/77.4.307
Calagari M, Modirrahmati AR, Asadi F (2006). Morphological variation in leaf trait of Populus euphratica Oliv.natural populations. International journal of Agriculture and Biology 8 (6): 754-758.
Casagrandi R, Gatto M (2002). Habitat destruction, environmental catastrophes, and metapopulation extinction. Theoretical Population Biology 61 (2): 127-140. https://doi.org/10.1006/tpbi.2001.1559
Cervera MT, Cervera VMT, Sewell MM, et al (2004). Genome mapping in Populus. In: Kumar S, Fladung M (eds) Molecular genetics and breeding of forest trees. Haworth’s Food Products, New York, pp 387-410.
Christerson L (2006). Biomass production of intensively grown poplar in the southernmost part of Sweden: Observations of characters, traits and growth potential. Biomass and Bioenergy 30 (6): 497-508. https://doi.org/10.1016/j.biombioe.2005.10.003
Dayanandan S, Dole J, Bawa K, Kesseli R (1999). Population structure delineated with microsatellite markers in fragmented populations of a tropical tree, Carapa guianensis (Meliaceae). Molecular Ecology 8 (10): 1585-1592. https://doi.org/10.1046/j.1365-294x.1999.00735.x
Dixon MD, Turner MG (2006). Simulated recruitment of riparian trees and shrubs under national and regulated flow regimes on the Wisconsin River, USA. River Research and Applications 22 (10): 1057-1083. https://doi.org/. 10.1002/rra.948
Fallah H, Tabari M, Azadfar D (2011). Determination ecotypes of Populus alba var. caspica Bornm. in plain communities of Caspian forests using morphological markers of leaf and peroxidase isoenzymes. Taxonomy and Biosystematics Journal. 3 (6): 47-59 (in Persian with an abstract in English). https://tbj.ui.ac.ir/article_17401.html?lang=en
Fallahchai M (2016). Natural regeneration of Populus caspica Bornm. and its relationship with soil physical and chemical properties (Case Study: Safrabaste region in east of Guilan province). Journal of Plant Research 29 (1): 118-129 (in Persian with an abstract in English). https://plant.ijbio.ir/article_913.html
Fang J (2012). Interspecific crossing breeding of Populus in Leuce section of China. In: Proceedings of the 24th Session of International Poplar Commission; India.
Gamari Zare A, Asadi F, Shariat A et al. (2020). The National botanical garden of Iran is a reliable source to supply healthy and high-vigor seeds of the native Populus caspica Bornm. Journal of Iran Nature 5: 49-57. (in Persian with an abstract in English). https://irannature.areeo.ac.ir/article_122532.html
Hamzeh M, Perinet P, Dayanandan S (2006). Genetic relationships among species of Populus (Salicaceae) based on nuclear genomic data. The Journal of the Torrey Botanical Society 133 (4): 519-527.
Hanski I, Ovaskainen O (2000). The metapopulation capacity of a fragmented landscape. Nature 404: 755-758. https://doi.org/10.1038/35008063
Homaie M, Mirzaie Nodoushan H, Asadicorom F, Bakhshi Khaniki GhR, Calagari M (2014). Evaluation of half-sib progenies and their parents of Populus euphratica based on their morphologic and micro-morphologic traits. Iranian Journal of Forest and Poplar Research 21 (4): 768-779 (in Persian with an abstract in English). https://doi.org/10.22092/IJFPR.2014.5149
Imbert E, Lefevre F (2003). Dispersal and gene flow of Populus nigra (Salicaceae) along a dynamic system. Journal of Ecology 91 (3): 447-456. https://doi.org/10.1046/j.1365-2745.2003.00772.x
Jafari Mofidabadi A (2005). Propogation of Populus caspica tree through mature ovary culture. Iranian Journal of Rangelands and Forests Plant Breeding and Genetic Research 13(1): 29-36 (in Persian). https://doi.org /10.22092/IJRFPBGR.2005.115214
Jalili A, Jamzad Z (1999). Red data book of Iran. Tehran, Iran: Research Institute of Forest and Rangelands (in Persian).
Khasa DP, Nadeem S, Thomas B, Robertson A, Bousquet J (2003). Application of SSR markers for parentage analysis of Populus clones. Forest Genetic 10 (4): 273-282.
Lopez DH, Sierra UR, Cristobal MD (2004). A comparison of isozyme and morphological markers to assess the within population variation in small populations of European aspen (Populus tremula L.) in Spain. Silvae Genetica 53 (5-6): 227-233. https://doi.org/10.1515/sg-2004-0041
Marron N, Dillen SY, Ceulemans R (2008). Evaluation of leaf traits for indirect selection of high yielding poplar hybrids. Environmental and Experimental Botany 61 (2): 103-116. https://doi.org/10.1016/j.envexpbot.2007.04.002
Rae AM, Robinson KM, Street NR, Taylor G (2004). Morphological and physiological traitsinfluencing biomass productivity in short rotation coppice culture. Canadian Journal of Forest Research 34 (7): 1488-1498. https://doi.org/10.1139/X04-033
Rechinger KH (1982). Flora Iranica: Akademische Druck-und Verlagsanstalt Graz. University of Tehran, p 549.
Sabeti H, 1976. Forests, trees and shrubs of Iran. Tehran, Iran: Agriculture and Natural Resources Research Organization Press (in Persian).
Sadati S, Tabari M, Assareh M, Heidari Sharifabad H, Fayaz P (2010). Effect of cutting source and planting depth on vegetative propagation of Populus caspica Bornm. Iranian Journal of Forest and Poplar Research 18 (4): 679-667. https://ijfpr.areeo.ac.ir/article_107684.html?lang=en
Storme V, Vanden Broeck AH, Ivens B, Smulders MJM, Halfmaerten D et al. (2002). Ex situ conservation of black poplar in Belgium, the margin of the geographical distribution area of the species In: Van Dam BC, Bordacs S, (editors). Genetic diversity in river populations of European black poplar. Budapest, Hungary: Csiszar Nyomda Ltd, pp 61-72.
Wan XQ, Zhang F, Zhong Y, et al (2013). Study of genetic relationships and phylogeny of the native Populus in Southwest China based on nucleotide sequences of chloroplast trnT–trnF and nuclear DNA. Plant Systematics and Evolution 299 (1): 57-65.
Yu Q, Tigerstedt PMA (2001). Growth and phenology of hybrid aspen clones (Populus tremula l. ã Populus tremuloides michx.). Silva Fennica 35 (1):15–25. https://doi.org/10.14214/sf.600
Young A, Boyle T, Brown AHD (1996). The population genetic consequences of habitat fragmentation for plants. Trends in Ecology and Evolution 11 (10): 413-418. https://doi.org/10.1016/0169-5347(96)10045-8
Yousefi B, Modir Rahmati AR (2004). Height growth behavior of poplar clones in Kurdistan. Iranian Journal of Forest and Poplar Research 13 (4): 533-556 (in Persian). https://ijfpr.areeo.ac.ir/article_109000.html?lang=en
Yousefzadeh H, Colagar AH, Yousefi E et al. (2019). Phylogenetic relationship and genetic differentiation of Populus caspica and Populus alba using cpDNA and ITS noncoding sequences. Journal of Forestry Research 30: 451-461. https://doi.org/10.1007/s11676-018-0785-4



